

26 Nov Más Veloz, Más Alto, Más Fuerte: ¿Qué Hay en el Menú?
Louise M. Burke1,2* and John A. Hawley2
1Australian Institute of Sport, Belconnen ACT 2616, Australia.
2Exercise and Nutrition Research Program, The Mary MacKillop Institute for Health Research, Australian Catholic University, Melbourne 3000, Australia.
*Correspondencia: louise.burke@ausport.gov.au
Las hazañas de los atletas élite nos deleitan, nos frustran y nos confunden a medida que se esfuerzan por alcanzar sus límites fisiológicos, psicológicos y biomecánicos. Aquí analizamos diversos enfoques nutricionales para un rendimiento óptimo, mostrando así la contribución que la ciencia deportiva moderna ha hecho en los títulos mundiales y las medallas de oro. A pesar de que aún se cree que existe una “dieta del atleta” única y superior, ha surgido la diversidad en las prácticas de nutrición deportiva entre atletas exitosos debido a la especificidad de las demandas metabólicas de los diferentes deportes y a la periodización del entrenamiento y de los objetivos de competencia. A esto se le debe añadir, la implementación pragmática de las estrategias de nutrición en escenarios de la vida real y la priorización de importantes estrategias cuando hay temas nutricionales en conflicto. Finalmente, las diferencias en las prácticas del atleta promueven y reflejan áreas de controversia y desacuerdo entre expertos nutricionistas del deporte.

Los atletas olímpicos antiguos manipulaban sus dietas de acuerdo a las creencias que predominaban en ese entonces, ganando crédito Pitágoras (probablemente de forma incorrecta) por haber iniciado a los atletas en el consumo de carne y de comidas ricas en proteína en lugar de los higos, cereales y quesos tradicionales (1). Entretanto, los atletas modernos son bombardeados por los “guerreros” de las redes sociales que evangelizan el veganismo, las dietas Paleo, y “keto” bajas en carbohidratos para mejorar el rendimiento. En contraste a la batalla del menú perfecto, la nutrición deportiva contemporánea apoya la diversidad en las prácticas dietarias, respaldando las demandas del entrenamiento y la competencia con las filosofías de especificidad, periodización y personalización (2). Las demandas metabólicas de los deportes élite son complejas, con eventos que duran desde segundos (saltos, lanzamientos y levantamientos) hasta semanas (carreras de ciclismo tipo Grand Tour). Los resultados en el desempeño se ven a partir del entrenamiento deliberado y específico de cada deporte el cual apunta a maximizar las adaptaciones para satisfacer el potencial individual genético (3). Aunque algunos atletas de élite se benefician de consejos sistemáticos y científicos con respecto a las adaptaciones al entrenamiento y al rendimiento en competencia, otros utilizan un enfoque de prueba y error bajo la guía de entrenadores experimentados, dejando así que sean los científicos los que expliquen post-hoc cómo la dieta pudo haber contribuido a sus picos de desempeño (3).
Resolviendo la Crisis Energética
La energía en los deportes competitivos es generada al transformar la energía química (glucógeno intramuscular y lípidos) en energía mecánica (contracción) siendo el trifosfato de adenosina (ATP) el intermediario (4). Debido a que las reservas intramusculares de ATP son pequeñas, los incrementos asociados al ejercicio en el movimiento de ATP dentro de los miocitos activos (hasta 100 veces la tasa de turnover en reposo) plantean un reto energético importante. Las vías metabólicas para re sintetizar ATP se activan de forma rápida durante sprints cortos (<30 segs), principalmente a través de la fosforilación a nivel de sustrato: la utilización de la fosfocreatina (PCr) y la conversión del glucógeno muscular en lactato. Sin embargo, a medida que la producción de ATP deja de igualar las tasas de utilización, se comienzan a acumular una variedad de subproductos. Algunos de ellos, como por ejemplo los iones de hidrógeno parecen ejercer una respuesta negativa en las vías que los producen para prevenir una perturbación en la homeostasis, mientras que otros, incluyendo el monofosfato de adenosina (AMP) y el fosfato orgánico, estimulan los sensores de energía tales como la 5’-proteína quinasa activada por AMP para mantener la homeostasis celular, regulando las vías anabólicas y catabólicas y asegurando, de ese modo, que exista un balance entre la oferta y la demanda energética.
Las actividades deportivas que duran desde varios minutos hasta varias horas y que se desarrollan en un estado continuo (p. ej., correr una maratón) o con arranques intermitentes a alta intensidad (p. ej., deportes de equipo) reciben su energía principalmente a través de la oxidación del glucógeno intramuscular y, en una menor medida, a través de los lípidos, mientras que la movilización de los sustratos extra musculares [glucosa plasmática del hígado y del intestino y ácidos grasos libres (FFAs) liberados por los adipocitos] se vuelve más importante a medida que la duración del ejercicio incrementa. El entrenamiento potencia la flexibilidad metabólica del miocito, mejorando el tamaño de las reservas de sustratos y la capacidad para cambiar rápidamente entre la energía proveniente de carbohidratos (CHO) y la proveniente de las grasas para poder satisfacer así las demandas de los músculos que se encuentran trabajando.
La ubicación – o el agotamiento específico en fibras de las reservas de glucógeno muscular se asocia con frecuencia con la fatiga, y desde la incorporación de la biopsia percutánea con aguja a la ciencia deportiva en los años 1960s (5), numerosas investigaciones han estudiado las estrategias para promover el almacenamiento del glucógeno antes o entre sesiones de entrenamiento. El factor clave en la síntesis de esta macromolécula es la cantidad de CHO dietarios consumidos (6), pero ya que las tasas máximas de síntesis por hora son equivalentes a ~5% del tamaño de las reservas normales, los atletas se ven en la necesidad de planear el momento adecuado, así como la ingesta suficiente de CHO para aumentar el glucógeno a niveles proporcionales a las demandas de la siguiente sesión (6). Los factores que reducen la eficiencia de la síntesis de glucógeno a partir de la ingesta de un CHO determinado incluyen daño muscular y el consumo inadecuado de energía, mientras que otros factores (estado de entrenamiento, severidad del agotamiento de glucógeno, consumo durante las horas siguientes a un ejercicio intenso y la co-ingesta de proteína) pueden incrementar las tasas de síntesis o las reservas absolutas (6). Las concentraciones en reposo del glucógeno en músculos entrenados para los deportes de resistencia son más altas que aquellas en individuos sedentarios. Además, la supercompensación de glucógeno (“carga de CHO”) se puede alcanzar, en músculos entrenados, en tan poco tiempo como 24 a 48 horas de actividad reducida y con altas ingestas de CHO [10 a 12g por kilogramo de masa corporal por 24 horas] (6). Durante al menos 50 años, estas técnicas han sido utilizadas por atletas para mejorar el rendimiento en eventos de resistencia donde el agotamiento de glucógeno ocurriría de no hacerse esto (7).
Por supuesto, las demandas de energía de muchos deportes exceden la capacidad de almacenamiento de glucógeno muscular y la oportunidad para el atleta de volver a llenar las reservas endógenas entre eventos. Los CHO exógenos (CHO consumidos durante las horas previas y/o a lo largo del ejercicio) mantienen la euglicemia ya que generan “un ahorro” en la producción de glucosa hepática (8), y la glucosa en sangre contribuye cada vez más a las tasas de oxidación de CHO muscular a medida que las reservas de glucógeno se agotan (9). Existen pruebas contundentes que sugieren que el desempeño en un rango de deportes y de escenarios deportivos es potenciado al consumir CHO durante el ejercicio, realizando ingestas orientadas hacia la necesidad que tiene el músculo de suplementar sus reservas decrecientes de glucógeno [30 a 60 g/hora en eventos de resistencia que duren hasta 2 o 3 horas y 60 a 90 g/hora en eventos de ultra distancia que duren entre 8 y 10 horas (10)]. La absorción intestinal de CHO es probablemente el peldaño limitante en la tasa de oxidación de los CHO ingeridos (11). Sin embargo, en cierta medida esto se puede solucionar “entrenando el intestino”: incrementando el consumo de CHO en la dieta y durante el ejercicio para aumentar la tolerancia y la actividad del transportador de glucosa dependiente del sodio SGLT-1 (12, 13). El uso de mezclas de glucosa-fructosa los cuales utilizan diferentes transportadores en el intestino también pueden incrementar la absorción intestinal total, así como las tasas de oxidación muscular de los CHO ingeridos (14).
Los almacenamientos relativamente grandes de lípidos aún en los atletas más magros tienen intrigados, con toda la razón, a los científicos deportivos quienes los ven como una fuente potencial de energía para ejercicios aeróbicos prolongados. No obstante, aunque la oxidación de CHO está estrechamente orientada a las demandas energéticas de los músculos que trabajan, no existen mecanismos que regulen de cerca la disponibilidad y el metabolismo de los FFAs para el gasto energético continuo. Las estrategias de corto plazo [ayuno nocturno o bajo en CHO, alimentación alta en grasa (LCHF)] no han demostrado ser exitosas en cuanto a mejorar el rendimiento se refiere, a pesar de incrementar la disponibilidad de FFA: El pequeño incremento en la oxidación de FFA es insuficiente para reemplazar la contribución de CHO después del agotamiento de las reservas de CHO musculares y hepáticas (15). Una estrategia alternativa (16) para potencialmente estimular el uso de ambas reservas de sustratos supone exponerse (5 días) a dietas bajas en carbohidratos y altas en grasas (LCHF por sus siglas en inglés) con 60% a 70% de grasas para promover el reequipamiento y mejorar el transporte y la utilización de FFA seguido por un reabastecimiento de la disponibilidad de CHO endógenos y exógenos (24 horas de una dieta alta en CHO y la ingesta de CHO antes y durante el ejercicio). A pesar de los incrementos sustanciales en las tasas de oxidación de grasa después de dichos protocolos, los beneficios en el rendimiento en actividades de resistencia han estado, en el mejor de los casos, limitados por los escenarios e individuos específicos (17).
En particular, la reducción observada en la utilización de CHO durante ejercicio submáximo (“ahorro de glucógeno”) la cual se consideraba inicialmente como favorable ya que preservaba esta energía para una oxidación tardía, se descubrió que era una oxidación deficiente de CHO causada por una glucogenólisis muscular reducida y por la regulación decreciente del flujo a través del ciclo ácido cítrico después de la reducción de la actividad de la enzima piruvato deshidrogenasa (PDH) (18). En deportes donde el éxito es determinado por un ejercicio aeróbico a altas intensidades ya sea a lo largo del evento (como en una prueba de ciclismo contrarreloj o en una carrera de 10.000 metros) o en etapas críticas [durante deportes en equipo o en los “arranques” y remates en las maratones, triatlones Ironman (19) y carreras más largas de ciclismo (20)] las tasas sostenibles más altas de turnover de energía muscular requieren de la mejor economía de producción de ATP a partir de la oxidación de CHO. Las estrategias a corto plazo de adaptación a las grasas, o incluso las adaptaciones a largo plazo de las dietas LCHF cetogénicas (80% grasa, <50g de CHO/día), las cuales pueden aumentar 2 o 3 veces las tasas normales de oxidación de grasa (21, 22) están limitadas en su aplicación a un pequeño rango de eventos deportivos, donde su utilización es lo suficientemente baja para que la energía del músculo sea proporcionada por la oxidación de grasa (21, 23). A la fecha, parece ser que los protocolos que aumentan significativamente la oxidación de grasas también disminuyen la flexibilidad metabólica al reducir las reservas de sustratos CHO y/o la habilidad para oxidarlos rápidamente. La conclusión es que cuando los atletas élite entrenan y compiten en la mayoría de eventos deportivos, los CHO son el sustrato predominante y crítico de los músculos que están trabajando y la disponibilidad de CHO (22, 24) en lugar de grasas, es la que gana las medallas de oro. Aquí proponemos que el incremento observado en las tasas de oxidación de grasas después de entrenamientos de resistencia y “entrenamientos con baja disponibilidad de CHO” (ver Cuando menos es más) es un indicador del incremento en la densidad mitocondrial; si se quiere tener éxito en las competencias, esta maquinaria será mejor utilizada si se aprovecha para mejorar la oxidación de la energía basada en CHO.
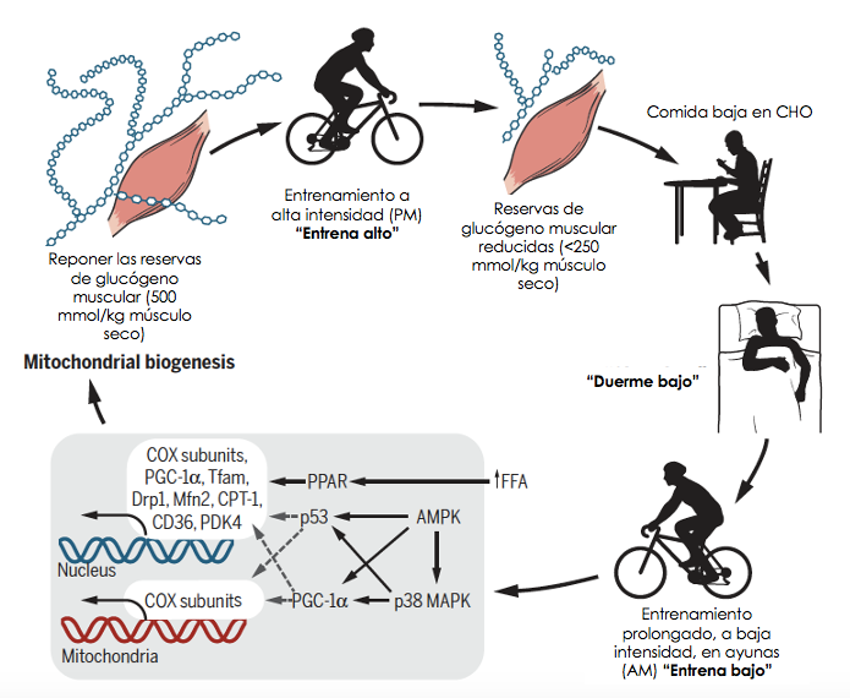
Figura 1. Nutrición periodizada: La evolución de una práctica nutricional. Iniciar un entrenamiento de resistencia con reservas bajas de glucógeno muscular (entrenar bajo) resulta en una mayor activación transcripcional de las enzimas involucradas en la oxidación de CHO y de grasas, así como en una mayor biogénesis mitocondrial comparado con hacer ejercicio con un contenido normal o elevado de glucógeno (29,30). Restringir la disponibilidad de CHO durante el período temprano de recuperación (1 a 5 horas) después del ejercicio también regula completamente varios marcadores del metabolismo de sustratos y la adaptación al entrenamiento de resistencia en el músculo esquelético (45). Utilizando estos antecedentes, formulamos un enfoque novedoso con el cual podemos realizar un entrenamiento de alta calidad y a alta intensidad y después prolongar la duración de la baja disponibilidad de CHO durante la recuperación y el subsecuente ejercicio aeróbico, extendiendo potencialmente la línea de tiempo de la activación transcripcional de los genes metabólicos y sus proteínas objetivo. Hemos llamado a esta práctica “entrena alto, duerme bajo” (45,46). PPAR, receptor activado por proliferadores de peroxisomas; AMPK, 5’-proteína quinasa activada por AMP; MAPK proteína quinasa activada por mitógeno; COX ciclooxigenasa.
Muévete, Músculo: ¡El Cerebro es el Jefe!
El cerebro y el SNC están implícitos en las tareas y en eventos que requieren de habilidad, concentración y toma de decisiones. Sin embargo, solo recientemente hemos podido reconocer su papel en el desempeño de eventos locomotores simples, incluidas las estrategias alrededor del ritmo de carrera. Hace un siglo, Bainbridge escribió, “Parece haber, no obstante, dos tipos de fatiga, uno que se genera en su totalidad en el sistema nervioso central, y otro donde la fatiga propia de los músculos se suma a aquella del sistema nervioso” (71). A pesar de esta perspectiva temprana, la nutrición deportiva ha evolucionado sesgada hacia el estudio de los mecanismos periféricos de la fatiga y su papel en el rendimiento, posiblemente por las oportunidades que ofrecen las herramientas disponibles de investigación (72). Se pueden observar excepciones; la hipoglicemia ha sido reconocida durante mucho tiempo como una causa de fatiga durante los deportes de resistencia (73), y hoy en día se sabe que los astrocitos del cerebro tienen reservas lábiles de glucógeno (74). Adicionalmente, ahora se pueden explicar los beneficios ergogénicos de la cafeína a través de los roles centrales (percepción reducida del esfuerzo y mayor reclutamiento neural de las fibras musculares) en lugar de su origen metabólico (ahorro del gluógeno muscular debido a una mayor disponibilidad y oxidación de los FFAs en plasma) (75). Un tema intrigante en la nutrición deportiva moderna involucra el SNC y los nutrientes que pueden mejorar el desempeño sin siquiera ser absorbidos (76). Este interés surgió a partir de la identificación de que la ingesta exógena de CHO podría mejorar el desempeño en ~1-hora de prueba de ciclismo de contrarreloj, aún cuando la disponibilidad del sustrato muscular (glucógeno) no sea limitada para este evento (76). Los mismos beneficios en el rendimiento se detectaron cuando la boca fue simplemente enjuagada con una solución de glucosa o maltodextrina (77), exponiendo los receptores de la cavidad bucal a los CHO y estimulando los centros de recompensa en el cerebro para aumentar el ritmo de carrera o el rendimiento en el entrenamiento (78). Se ha notado que los beneficios de la “sensación oral” de los CHO son robustos y repetibles cuando se llevan a cabo durante un evento, ofreciendo nuevas estrategias de nutrición para el rendimiento y un rango diferente de los deportes a los que orientan estas estrategias (76). Aunque esta ciencia aún se encuentra en sus primeros pasos, la evidencia muestra los beneficios de las sensaciones orales a partir de otros gustos (p. ej., fluidos y cafeína), y se han reportado efectos para el mentol (percepción de frescura), la quinina [activación del sistema nervioso autónomo y/o la excitabilidad corticomotora para eventos breves (79)], y la capsaicina, el ácido acético, el cinamaldehído, y otros químicos de las plantas que son conocidos activadores de canales receptores y que podrían prevenir los calambres musculares asociados al ejercicio (80).
Entrenando la Nutrición: Un Acto de Balance
Una visión reduccionista del entrenamiento, identifica una variedad de adaptaciones interdependientes que permite a los atletas sostener la tasa y rentabilidad más altas de la producción de energía, optimizar la economía del movimiento, defender la homeostasis celular, y retrasar el inicio de la fatiga al mismo tiempo que se alcanza el rendimiento físico óptimo y las habilidades técnicas específicas de sus eventos (3, 25). Para lograr estos fines, los atletas élite y sus entrenadores integran una serie de entrenamientos que individualmente abordan atributos importantes del rendimiento en competencia, y los incluyen dentro de un programa de entrenamiento periodizado compuesto por microciclos (semanales) y mesociclos (3-6 semanas), culminando en picos específicos de competencia dentro del macro ciclo (anual). Las adaptaciones del entrenamiento a largo plazo, como las observadas en atletas élite, resultan por el efecto acumulado de varios incrementos transitorios en las transcripciones de mRNA que codifican varias proteínas después de cada sesión intensa de entrenamiento (26, 27). Estas ráfagas repetidas en las expresiones de mRNA parecen ser esenciales para accionar la respuesta adaptativa intracelular crónica del entrenamiento (27).
Con respecto a la dieta del entrenamiento, las primeras guías nutricionales promovían un enfoque único y de cierta forma estático, concentrándose predominantemente en “combustible” basado en CHO como la principal fuente de energía para el músculo. Sin embargo, las guías contemporáneas reconocen que existen diferencias en los requerimientos y objetivos de las diversas sesiones o fases del entrenamiento, lo cual ha conducido a la periodización de la dieta del atleta (2, 28). En principio, el apoyo nutricional está organizado alrededor de cada sesión de entrenamiento para maximizar los resultados fisiológicos dentro de un marco que tenga en cuenta las metas nutricionales más grandes. Las recomendaciones tienen como objetivo la energía total y la disponibilidad de “combustible”, con un foco en la ingesta de proteína y de CHO y su distribución durante el día (2). Para las sesiones de alta intensidad o alta calidad, se recomienda tener una gran disponibilidad de CHO endógenos y exógenos, de tal forma que la programación en el tiempo y la ingesta de CHO coincidan o excedan los requerimientos energéticos del músculo respaldando así la calidad del entrenamiento (2, 28, 29). Para algunas sesiones de entrenamiento se deberían desarrollar planes de ingesta de CHO y fluidos para competencia, incorporando la práctica de los comportamientos específicos del evento, así como entrenando la función intestinal que se necesita para tolerar y liberar estos nutrientes al flujo sanguíneo. La ingesta de CHO puede ser reducida para cargas de entrenamiento más livianas (2, 29); asimismo, las adaptaciones al entrenamiento de resistencia pueden ser mejoradas al iniciar algunas sesiones, de forma deliberada, en condiciones de baja disponibilidad de CHO endógenos y exógenos (28-30) (ver Cuando menos es más y la Fig. 1.). Como resultado, las ingestas diarias de CHO variarán típicamente entre 2 y 12 gr por kilogramo masa corporal entre atletas y entre ciclos de entrenamiento (31).
Las nuevas perspectivas sobre la ingesta de proteína apuntan al consumo diario de cantidades útiles (~0.3 g por kilogramo de masa corporal) que provengan de fuentes de alta calidad repartidas en 4 o 6 comidas o snacks, particularmente durante la ventana entre 1 y 3 horas después de sesiones clave de entrenamiento (32, 33) y, tal vez, una dosis doble antes de dormir (34). Este enfoque amplía el argumento ya existente de que los atletas tienen requerimientos diarios de proteína por encima del consumo diario recomendado estándar, que necesitan para optimizar el incremento sostenido (~24 horas) en la síntesis de la proteína muscular estimulada por contracción y que ocurre después de un ejercicio de fuerza o de resistencia al proporcionar leucina que regule la vía de la diana de rapamicina en células de mamífero (mTORC1) (32,33). De igual forma, es importante prestar atención a los nutrientes para los cuales los atletas tienen un requerimiento mayor [como el agua, los electrolitos, y el hierro (35)] o para los cuales tienen riesgo de sufrir deficiencias [como la vitamina D (36] y que juegan un papel importante en maximizar las adaptaciones al entrenamiento, especialmente durante sesiones que se llevan a cabo en el calor o en altitudes elevadas (3).
Los atletas élite se mueven en una delgada línea entre maximizar el estímulo general del entrenamiento para promover las adaptaciones específicas del deporte y permanecer libres de enfermedad o lesión. Dentro del marco de cualquier programa de entrenamiento periodizado, cada sesión intensa de ejercicio hace parte integral de una meta a largo plazo, tratando de balancear finamente el impulso del entrenamiento (la suma de la intensidad, la duración, y la frecuencia de las sesiones) y así afianzar adaptaciones óptimas para un pico específico de competencia. El integrar metas nutricionales en una sesión o fase específica dentro de un plan nutricional más amplio crea con frecuencia tensión entre temas opuestos. Los atletas, tanto hombres como mujeres, pueden desarrollar una deficiencia de energía relativa en el deporte (RED-S por sus siglas en inglés, relative energy deficiency in sport) debido al desequilibrio que se crea entre la energía que se consume y la energía que se gasta en el ejercicio, lo cual conlleva a alteraciones en la tasa metabólica, la salud ósea, la síntesis de proteínas, la producción de hormonas reproductivas y las ganancias en el rendimiento (37)

Sin embargo, el RED-S puede ocurrir de forma secundaria a la manipulación de la grasa corporal y a los muy altos volúmenes de entrenamiento que típicamente aseguran el éxito (38); esto necesita de una implementación cuidadosa y de una programación de dichas fases en el plan anual (39). Otras estrategias que necesitan ser balanceadas o manejadas de forma estratégica incluyen el entrenamiento con disponibilidades bajas de CHO, el cual incrementa la respuesta inflamatoria (interleucina-6) del ejercicio, con grandes perturbaciones al sistema inmunológico (40), y al metabolismo óseo (41) el cual parece persistir por la exposición a largo plazo (42). De forma similar, el uso de suplementos antioxidantes reduce de manera importante el daño de los radicales libres de oxígeno a las fibras musculares, pero perjudican potencialmente la respuesta al entrenamiento al interferir con las vías de señalización sensibles al estado redox (reducción-oxidación) esto se ha traducido a su vez en una disminución en el rendimiento (43).
Cuando Menos es Más
La aplicación de técnicas de biología molecular a la ciencia del ejercicio ha identificado la complejidad y el alcance de las redes de señalización intracelular mediante las cuales las diversas formas de ejercicio impulsan los cambios adaptativos que respaldan el fenotipo atlético (3, 25). El alterar la disponibilidad de nutrientes, particularmente las reservas de CHO endógenos, modula selectivamente la expresión génica y la señalización intracelular dentro del músculo; los mecanismos incluyen alteraciones en la osmolaridad de la célula y una mayor actividad de las moléculas dentro del dominio regulador del CHO-vinculante de la proteína quinasa activada por AMP, así como perturbaciones a los FFAs y las hormonas que circulan en colaboración con la glucosa plasmática y las concentraciones de insulina (3, 29). Dentro del repertorio de las estrategias de nutrición para el entrenamiento, los atletas ahora pueden incluir prácticas que aumenten los procesos adaptativos en el músculo esquelético; estos incluyen comenzar a entrenar con una disponibilidad baja de CHO exógenos (ayuno nocturno y/o suspender el consumo de CHO durante una sesión) o la potente estrategia de entrenar bajo donde deliberadamente se comienzan determinadas sesiones con reservas reducidas de glucógeno muscular (p.ej., utilizar una primera sesión para agotar el glucógeno y entrenar una segunda vez después de privarse de consumir CHO para prevenir el restablecimiento de glucógeno) (29, 30).
Aunque algunos estudios reportan de forma consistente sobre el incremento en las respuestas celulares como resultado de las estrategias de entrenamiento bajo, su traducción en la mejora de rendimiento no ha sido tan clara (29, 30). Las investigaciones tempranas han fallado en detectar resultados superiores en el rendimiento; esto ha sido atribuido al excesivo énfasis de dichas sesiones dentro del programa de entrenamiento y su consecuente afectación en la intensidad del mismo (44). Estas sesiones deben ser ubicadas apropiadamente dentro de un programa periodizado para complementar un entrenamiento de alta calidad (7). Una reciente e ingeniosa secuencia de prácticas (Fig. 1) integra una sesión que promueve el rendimiento y una sesión enfocada en adaptación al tiempo que añade los beneficios de un incremento prolongado en la señalización celular estimulada por el ejercicio así como en la regulación post-transcripcional durante el ejercicio y la recuperación del glucógeno agotado (45). En poblaciones sub-élite al menos, una mejor integración entre sesiones de entrenamientos bajos y entrenamientos altos dentro de la secuencia de entrenamiento (Fig. 1) ha sido asociada con un rendimiento superior comparado con llevar a cabo el mismo entrenamiento pero con disponibilidades normales de CHO (46). Sin embargo, hasta la fecha, este no parece ser el caso en estudios que involucran poblaciones élite (22, 47), a pesar de ser incorporados con frecuencia en sesiones de entrenamiento reales (48). Aunque se necesitan aún más estudios, una parte del reto que existe para avanzar en esta área de investigación es que no se han llegado a acuerdos con respecto a la terminología e implementación de las prácticas involucradas; hemos tratado de abordar este tema en un comentario separado (7).
Rendimiento en Botella
Los productos deportivos representan una porción lucrativa de la explosión mundial en la fabricación y mercadeo de suplementos; de acuerdo con un reporte, los suplementos deportivos generaron ingresos globales de $9 billones en 2017, con el pronóstico de doblar este número para 2025 (81). Las encuestas confirman que el uso de alimentos y suplementos deportivos tienen una alta prevalencia entre atletas, incluyendo un mayor uso a medida que los niveles de competencia son más altos (82). A pesar de existir una temprana reticencia, numerosos grupos de expertos, incluyendo el Comité Olímpico Internacional (83) ahora acepta pragmáticamente el uso de suplementos después de que un análisis de riesgo-beneficio mostrara que es seguro, efectivo, legal y apropiado para la maduración del atleta en su deporte. Los suplementos utilizados por los atletas se encuentran en diferentes categorías (83): suplementos nutricionales para el tratamiento o prevención de deficiencias (p.ej., hierro y vitamina D); alimentos deportivos que proporcionan energía o nutrientes cuando no es práctico consumir comida “normal” (p. ej., bebidas deportivas y suplementos de proteína); suplementos para el rendimiento que mejoran directamente la capacidad del ejercicio; y suplementos que proporcionan beneficios indirectos a través de la recuperación, manejo de la composición corporal, entre otros objetivos. A pesar de la entusiasta publicidad, de los dos últimos grupos, solo unos pocos productos gozan de robusta evidencia acerca de su eficacia [p. ej., cafeína, monohidrato de creatina, bicarbonato, beta-alanina, y nitrato (84)] (Fig. 2).
Cualquier beneficio asociado al uso de suplementos debe ser medido contra el gasto, el potencial de sufrir efectos adversos debido a los pobres protocolos de utilización (p. ej., dosis excesivas o interacción con otros suplementos) y los peligros inherentes a los productos cuya manufactura y mercadeo son menos regulados que aquellos para los alimentos o bienes farmacéuticos. Existen varios asuntos de seguridad alrededor de los suplementos dietarios en general los cuales representaron ~23.000 visitas reportadas en el servicio de urgencias en Estados Unidos en el 2015 (85). Los atletas élite también necesitan considerar que se ha encontrado que los suplementos contienen contaminantes o ingredientes no declarados que están prohibidos por los códigos antidoping (86) bajo los cuales compiten; estos incluyen estimulantes, agentes anabólicos, moduladores selectivos del receptor de andrógeno, diuréticos, anorexígenos y agonistas b2 (87). Los estrictos códigos de responsabilidad suponen que un test de orina positivo puede desencadenar en una violación de las reglas antidopaje con efectos potenciales serios en la carrera del atleta, sus medios de subsistencia, y reputación a pesar de ser una ingesta no intencional o de dosis minúsculas (inefectivas). Las auditorías de producto a manos de terceros pueden ayudar a los atletas élite a tomar decisiones informadas sobre el uso de suplementos más no podrán ofrecer una garantía absoluta acerca de la seguridad del mismo (83).
Luchando contra la Fatiga: Comer para Ganar
Cada deporte tiene características distintivas, pero las características que se comparten entre todos los competidores son el deseo de marcar el ritmo de su rendimiento para lograr los resultados o velocidades más altas que se puedan sostener y mantener una proficiencia técnica, con la posibilidad de que se reduzcan algunas métricas de desempeño de forma intermitente hacia el final del evento, o que sucedan ambas cosas. La “fatiga” se define operacionalmente como un descenso periódico o sostenido en la habilidad del atleta para desempeñar de forma óptima las tareas requeridas por su deporte. Aunque la fatiga se caracteriza con frecuencia como muscular (producción de fuerza o potencia reducida) o mental (mayores niveles de percepción de esfuerzo o pérdida de las habilidades cognitivas) existe una interacción entre ambos fenómenos. La fatiga muscular tiene entradas tanto periféricas (relacionadas con el músculo que se ejercita) como centrales [relacionadas con la habilidad del sistema nervioso central (SNC) para enervar las fibras musculares]. Aunque algunos eventos requieren de un rendimiento máximo para romper marcas mundiales o personales, otros premian un rendimiento superior relativo al de los otros competidores. Los factores que promueven la fatiga o el rendimiento en términos de potencia o velocidad son específicos para cada evento, el entorno en el que se desarrolle y la individualidad del atleta.
La Figura 2 muestra un resumen simplificado de los factores de fatiga más comunes en deportes de competencia a los cuales se les puede aplicar las estrategias de nutrición basadas en evidencia para reducir o retrasar el inicio de la fatiga. Dichas estrategias pueden involucrar protocolos crónicos que trabajen en sinergia con el entrenamiento para volver al cuerpo más resiliente a estos factores. Por ejemplo, en los deportes de equipo, los cuales implican la repetición de sprints cortos (p.ej., 6 segs) y de alta intensidad, el deterioro progresivo de la velocidad que está relacionado con la inhabilidad de recuperar por completo las reservas de PCr en los periodos de recuperación (>30 a 120 s) se puede solucionar a través de la suplementación con monohidrato de creatina para incrementar el tamaño de las reservas musculares de PCr (49). Esto podría no solo mejorar directamente el desempeño en los partidos al alterar el deterioro del perfil de sprint (50) sino que también le permitiría al atleta entrenar más duro (p. ej., completar más sprints durante las sesiones de entrenamiento) para incrementar las adaptaciones en otros sistemas fisiológicos. Las estrategias intensas previas al evento tales como la carga de CHO (24 a 48 horas de preparación) para aumentar las reservas de glucógeno muscular o la hiper-hidratación asistida por el glicerol (2 horas de preparación) para elevar los depósitos de agua corporal pueden potenciar el rendimiento en eventos específicos si logran incrementar el tiempo de potencia óptima antes de que el cuerpo alcance un nivel crítico de agotamiento de glucógeno (51) o un déficit de fluidos (52), respectivamente. La ingesta de CHO (10) o fluidos (53) durante el evento pueden solucionar también estos factores limitantes secundarios pero, misteriosamente, pueden proporcionar beneficios a través de un efecto del SNC asociado con la sensación oral de estos nutrientes.
Finalmente, para eventos de la vida real, el desarrollo de un plan de nutrición de competencia está desafiado por la complejidad de pensar en una multitud y por la superposición de los factores de fatiga, por las limitaciones prácticas impuestas por el evento o por la naturaleza de la actividad, y por las creencias y la tolerancia del atleta. Esto se puede ver relejado en el Cuadro 3 y es ilustrado por actividades recientes en maratón (42.2km). En 2017, en un intento cuidadosamente orquestado para romper la barrera de las 2 horas (54) y después de muchas bromas científicas (55) el keniata Eliud Kipchoge llegó 25 segundos por encima de las 2 horas. Kipchoge contaba ya con dos de los tres “factores de éxito” para eventos largos de resistencia: una potencia aeróbica alta y la habilidad para correr a una muy alta proporción de su capacidad aeróbica por periodos prolongados sin perder el control metabólico (56). Su intento, aunque por fuera de las reglas de la Asociación Internacional de Federaciones de Atletismo, le apuntó principalmente al tercer factor: la economía de carrera (alcanzar la más alta velocidad por el menor costo de oxígeno). Las estrategias para mejorar la economía de carrera incluyeron correr en una pista plana de carreras automovilísticas sin esquinas agudas para conservar la velocidad, correr en una formación aerodinámica detrás de otros corredores, utilizar un cronómetro montado en un carro para ofrecer un efecto parabrisas así como para asistir con la marcación del ritmo y el uso de zapatillas diseñadas para devolver un 4% de energía extra a través de insertos de fibra de carbono (57).
También se utilizaron estrategias nutricionales para optimizar la economía, y es muy probable que haya futuras mejoras. Los efectos beneficiosos del jugo de remolacha, una ayuda popular para el rendimiento que proporciona una fuente adicional de nitratos inorgánicos, están mediados a través de la mejora de la economía del ejercicio: Se cree que una vía generadora de (NO) óxido nítrico (nitrato-nitrito-NO) mejora los incrementos mediados por el NO en la distribución de O2 capilar al músculo y reduce las fugas de protones mitocondriales (58). Además, a pesar de las afirmaciones actuales de que las dietas cetogénicas LCHF proporcionan sustratos ilimitados para las actividades físicas prolongadas (21), tanto los datos empíricos de hace más de un siglo (59) como las intervenciones recientes que involucran senderistas de competencia de talla mundial (22) nos recuerdan que la oxidación de CHO produce ~5% más ATP por unidad de O2 que la grasa. Las estrategias de nutrición del futuro para maratón se podrán enfocar en una mayor disponibilidad y oxidación de CHO alejándose del uso de la grasa que proviene del horno mitocondrial. Algunas tácticas incluyen alcanzar reservas máximas de glucógeno supercompensadas, aumentar las oportunidades de consumo de alimento durante la carrera, y entrenar el intestino para utilizar múltiples fuentes de CHO incrementando así la absorción total de CHO (12).
La Especificidad y Practicidad Requieren de Soluciones Personalizadas
La implementación práctica de las estrategias de nutrición hecha por atletas en escenarios reales confunde el establecimiento de una base de evidencia por métodos tradicionales de investigación y el desarrollo de guías que se puedan generalizar (y que no sean controvertidas). En la mayoría de los deportes el rendimiento está limitado por un número factores independientes, que típicamente se abordan a través un plan coordinado. Frecuentemente consideramos una combinación de varias estrategias de nutrición independientemente valiosas, a pesar de que sus efectos puedan ser potencialmente redundantes, amplificadores, atenuantes o que compitan entre sí. Los beneficios individuales de la reposición de fluidos y de CHO para el desempeño en el calor son aditivos (88), la cafeína es menos efectiva cuando la ingesta de CHO es también utilizada para atenuar la disminución del rendimiento durante ejercicios prolongados (89), y combinar la cafeína con una suplementación de bicarbonato afecta los beneficios de la primera debido a perturbaciones gastrointestinales (90). Sin embargo, debido a que es poco práctico investigar todas las permutaciones y combinaciones de las estrategias de nutrición basadas en evidencia (90), los efectos generales en el rendimiento son desconocidos.
Las condiciones medio ambientales y el calendario de competencias añaden aún más retos prácticos. Los campeonatos Premiere pueden ser organizados bajo condiciones “hostiles” (como la elevada altitud en los Juegos Olímpicos de Ciudad de México 1968 o el calor del Campeonato Mundial de Atletismo en Doha 2019) o en horarios inusuales (como las competencias de natación tarde en la noche en los Juegos Olímpicos de Rio 2016 que se llevaron a cabo a esa hora para que coincidieran con las horas de mayor audiencia de la televisión de Estados Unidos). De este modo, los atletas necesitaron con frecuencia de estrategias personalizadas para diferentes escenarios del mismo evento. Finalmente, los deportes suponen reglas, consideraciones logísticas y culturas que dictan oportunidades para la ingesta de nutrientes antes, durante y entre eventos (91). Algunos ofrecen oportunidades adecuadas para llevar a cabo una ingesta que sea beneficiosa (p.ej., los jugadores de basquetbol toman líquidos durante las sustituciones y los tiempos de descanso). Sin embargo, las condiciones en otros eventos, como los deportes de la división de peso, fomentan una deshidratación sustancial antes del evento, y una restricción de energía para “hacer peso” (92). El fútbol prohíbe los descansos o el acceso a fluidos durante cada mitad de juego y el reto práctico de tomar líquidos mientras se corre a ~21 km/hora en una maratón limita el volumen que se ingiere (68).
Es importante considerar si la conducta tradicional y la evaluación de la investigación científica informa adecuadamente a los atletas élite. En muchas áreas de nuestras vidas, nos sentimos satisfechos con las verdades (la estrategia X es buena) y las guías (todos deberíamos implementar la estrategia X haciendo Y) que se pueden generalizar. La Figura 3 ilustra la jerarquía de los tipos de evidencia científica. Los tipos de estudios que proporcionan la evidencia más fuerte o de más alta calidad (como los ensayos controlados aleatorios) son extremadamente difíciles de conseguir con competidores de alto calibre o pueden proporcionar información genérica inapropiada para una tarea específica. La falta de apreciación de estos conceptos ha causado angustia entre la comunidad de ciencia deportiva y un rechazo injusto sobre la integridad de sus resultados.
La ciencia deportiva fue criticada en una evaluación hecha por científicos entrenados en epidemiología (93,94). Aunque se plantearon cuestiones metodológicas válidas, el análisis no logró apreciar que los científicos deportivos que trabajan en deportes élite típicamente buscan información altamente específica al contexto (Fig. 3). Las soluciones basadas en evidencia, aunque personalizadas a un pequeño número de individuos, requieren de diseños y herramientas de investigación especiales; la validez, fiabilidad y sensibilidad de las medidas son críticas. Debemos considerar la reproducibilidad en el diseño del estudio o en los enfoques de la historia clínica para justificar el tamaño pequeño de las muestras. Incluso la capacidad de respuesta individual a las intervenciones puede variar. Es posible que se requiera de enfoques estadísticos nuevos o más refinados (60, 90).
La controversia que gira en torno a las guías de ingesta de fluidos durante la práctica deportiva ejemplifica la falta de comprensión que existe alrededor del contexto. Los críticos de las guías actuales que apoyan un enfoque individualizado (95) y quienes sostienen que a los atletas se les debería decir que consuman líquidos solo cuando sientan sed durante los eventos (96) fallan en reconocer que las oportunidades para ingerir fluidos están frecuentemente fuera del control del atleta y no están sujetas a la necesidad. Por lo tanto, es razonable desarrollar un plan personalizado para eventos específicos que optimice la oportunidades para consumir fluidos y CHO antes y durante el evento y donde se integren la comodidad del intestino, las necesidades energéticas, un déficit de fluidos tolerable y el manejo de la sed.
Los Atletas de Élite son Diferentes
El escrutinio de la evidencia base para las guías actuales de nutrición deportiva revela que los individuos que contribuyen a las investigaciones científicas con sangre, sudor y lágrimas están a lo sumo bien entrenados, son frecuentemente hombres y casi siempre sub-élite. Las intervenciones hechas en atletas de talla mundial son inusuales: Por definición, dichos atletas son escasos en número y por lo general no están inclinados a interrumpir un entrenamiento o plan de nutrición exitosos o someterse a técnicas experimentales invasivas solo por el beneficio de la ciencia. Por lo tanto, es razonable preguntar si los resultados de los estudios hechos en poblaciones no-élite aplican también a sus contrapartes élite. Algunos temas incluyen la aplicación de la intervención a escenarios específicos en los que los atletas élite compiten o entrenan, la inhabilidad de los estudios para detectar diferencias o cambios importantes aunque pequeños con relación al rendimiento y que podrían alterar los resultados de un deporte élite y la traducción de los mecanismos putativos a los atletas que llevan a cabo volúmenes de entrenamiento especializado sustancialmente más altos y que poseen potencialmente rasgos genéticos favorables (60).
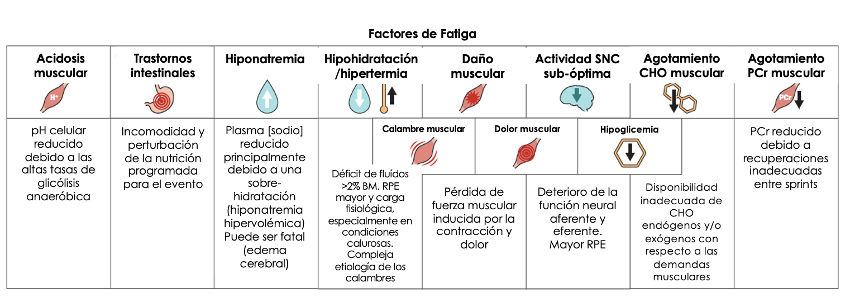
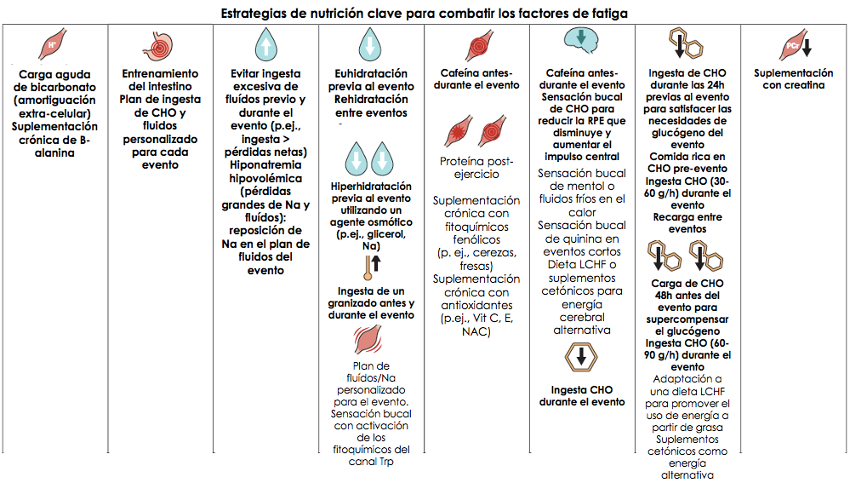

Figura 2. La nutrición puede combatir la fatiga de la competencia. Muchos factores que comúnmente causan fatiga (una disminución periódica o sostenida en la habilidad del atleta para desempeñarse de manera óptima) en eventos deportivos se pueden abordar con estrategias nutricionales que reduzcan los efectos de estos factores o que retrasen su inicio.
En relación con algunas intervenciones de nutrición, la evidencia sugiere que hay una respuesta reducida en competidores élite. Por ejemplo, la suplementación con jugo de remolacha parece ser menos efectiva a la hora de lograr mejoras en la economía o en el rendimiento de atletas élite (61); las explicaciones para esta falta de respuesta de parte de dicha suplementación en atletas de alto calibre incluyen su composición muscular diferente y el legado de las adaptaciones fisiológicas logrado a través de un entrenamiento extensivo, tal como un mayor grado de actividad de la vía principal NO-arginina (62, 63). Sin embargo, ya que esta vía es dependiente del oxígeno, se podrían identificar escenarios donde los atletas élite se beneficien de una producción alternativa (independiente de oxígeno y pH) de NO derivado de nitratos, justificando así la suplementación con jugo de remolacha; estos incluyen entrenar o competir en altitudes elevadas y competir en deportes que involucren grupos musculares pequeños, como los brazos, en los cuales el bajo flujo sanguíneo aumenta la posibilidad de sufrir de hipoxia local y acidosis (62, 63).

El reto sigue siendo determinar si los atletas élite son exitosos gracias a, o a pesar de, sus prácticas nutricionales. Existen pocos estudios de dichos grupos o individuos, aunque los atletas élite de África oriental quienes han dominado las medias distancias en running durante las últimas décadas han recibido investigación científica (64,65). Sus patrones alimenticios incluyen consistencias con las guías actuales del atleta [ingestas altas de CHO (~60 al 80% de la energía) pero con uso regular de los entrenamientos en estado de ayuno para lograr las sesiones de entrenamiento bajo], así como inconsistencias ya sea con las guías o con las prácticas típicas de otros atletas élite [dependencia en las fuentes de origen vegetal (80 a 90% de la dieta) en lugar de animal, variedad de comida muy limitada, distribución de energía en una pequeña cantidad de comidas al día, y periodos crónicos de disponibilidad baja de energía]. Algunas historias clínicas con enfoques concebidos por científicos para el rendimiento de competidores élite, tales como la “fabricación de peso” para un boxeador profesional (66) o el complejo plan de nutrición seguido por el ganador de la competencia ciclística de 3 semanas, el Giro d’Italia, no necesariamente permiten sacar conclusiones firmes y replicables sobre los beneficios de estas estrategias pero sí muestran cómo ciertas prácticas se pueden lograr dentro de las complejidades del entorno deportivo (ver Fig. 3).
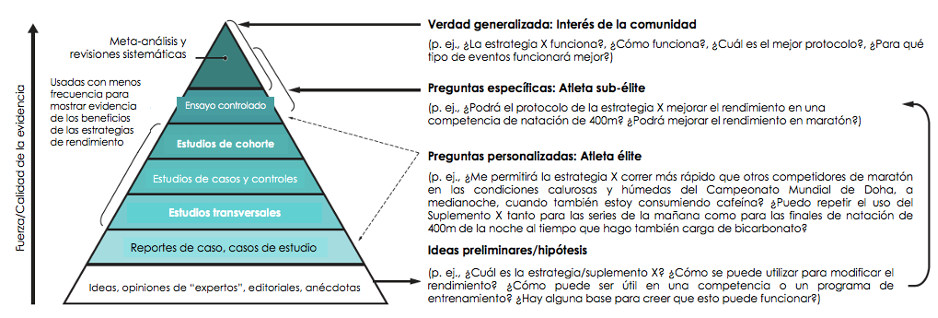
Figura 3. Perspectivas sobre la base de evidencia para las prácticas de atletas élite. Desarrollar una base de evidencia para las prácticas nutricionales de los atletas élite requiere reconocer que se necesitan respuestas específicas a las preguntas de investigación e interpretaciones de las guías existentes.
En el último caso, el plan de competencia manipuló la masa corporal (al restringir ligeramente la energía y usar una dieta baja en residuos para reducir los contenidos gastrointestinales) de acuerdo a las ventajas de ser más ligero en las secciones de montaña y fluctuó la ingesta diaria de energía y CHO de acuerdo a los costos metabólicos estimados de cada etapa (67). Vale la pena notar, que el día decisivo de esta carrera, el cual involucraba un recorrido a solas por un terreno montañoso de alta dificultad, se logró con un plan de nutrición que proporcionaba 6663 kcal y 18.9 g de CHO por kilogramo (un total de 1.3 kg, equivalentes a ~85 tajadas de pan) y con las provisiones de carrera siendo proporcionadas por los compañeros de equipo y el equipo de apoyo en intervalos programados para evitar que el ciclista tuviera que cargar con el peso (67). Nunca sabremos cuánto contribuyó este plan al eventual éxito del ciclista. Igualmente, necesitamos racionalizar que el éxito atlético se puede lograr de cara a una aparente práctica por debajo de niveles óptimos. Por ejemplo, que el ganador de una maratón élite haya incurrido en una pérdida del 10% de la mas coporal a lo largo de la carrera (68) falla en refutar que la hipo-hidratación perjudica el rendimiento; en su lugar, demuestra que este atleta fue más rápido que otros competidores ese día y que potencialmente fue el que mejor toleró las condiciones.
Si bien los atletas élite pueden aprender de la ciencia deportiva, muchas lecciones también han corrido en la dirección opuesta. Las recomendaciones de nutrición deportiva son a menudo actualizadas cuando las prácticas observadas entre atletas élite parecen ser beneficiosas. Por ejemplo, las guías de la cafeína para el deporte cambiaron cuando se descubrió que las bebidas de cola consumidas por ciclistas élite hacia el final de las carreras largas (~1 a 2 mg de cafeína por kilogramo al inicio de la fatiga) eran tan efectivas como los protocolos “científicamente comprobados” (6 a 9 mg/kg ingeridos 1 hora antes del evento) (69). La munición para actualizar las guías de ingesta de CHO durante eventos prolongados (>2.5 horas) se generó al observar que la ingesta de varios ciclistas y triatletas élite (~90 g/hora) fue mayor a la que se recomendaba antes (30 a 60 g/hora) y tenía correlación con el éxito (70). Claramente, los resultados futuros se alcanzarán de una mejor manera cuando haya una interacción de doble vía entre los científicos deportivos y los atletas élite y sus entrenadores. En los análisis finales, la nutrición deportiva moderna ofrece un banquete de oportunidades para asistir a los atletas élite para que entrenen duro, optimicen las adaptaciones, estén saludables y libres de lesiones, para que logren la composición corporal deseada y para que puedan luchar contra los factores de fatiga que limitan el éxito. Aunque habrá retos y cambios en las guías de nutrición deportiva a medida que evolucionan más allá de las fronteras del conocimiento y la práctica actual, podemos emocionarnos al saber que la ciencia deportiva en muchos de sus aspectos contribuye a los resultados que nos deleitan y nos sorprenden desde nuestros sofás y desde las tribunas.

Traducción
Juliana Bermeo
Referencia Original
Burke Louise M. and John A. Hawley. Swifter, higher, stronger: What’s on the menu?
Science 362 (6416), 781-787. DOI: 10.1126/science.aau2093.
Reconocimientos
Intereses opuestos: L.M.B es un director del programa del International Olympic Committee Diploma in Sports Nutrition y ha sido financiado por el Australian Institute of Sport High Performance Research Funds, Australian Catholic University, y el Australian Research Council, así como por colegas de la industria que incluyen el Alliance for Potato Research and Education (APRE), South African Potato Producers Organisation, Dairy Health and Nutrition Research Consortium, Dairy Australia, Nestle Research Centre, Nestle Australia, Kellogg’s Australia, Mars Australia, and Gatorade Australia. L.M.B. fue miembro del Panel de Expertos del Gatorade Sports Science de 2014 a 2015 por cuyo trabajo recibió honorarios. J.A.H. ha recibido financiación para realizar estudios de metabolismo nutricional por parte de APRE; el Novo Nordisk Foundation; el Australian Sports Commission; Dairy Health and Nutrition Research Consortium, Australia; el Australian Research Council; Nestec, Switzerland; the National Heart Foundation of Australia; the Diabetes Australia Research Trust; Uncle Ben’s of Australia, una division de EFFEM Foods; Polar Electro, Finland; SmithKline Beecham Consumer Healthcare (Nutrition), United Kingdom; the South African Potato Board; Bromor Foods, South Africa; the Sugar Association of South Africa; and Wander Research and Development, Switzerland. J.A.H. fue miembro del Panel de Expertos del Gatorade Sports Science de 2014 a 2015.
Referencias y Notas
- L. E. Grivetti, E. A. Applegate, J. Nutr. 127 (Suppl), 860S–868S (1997).
- D. T. Thomas, K. A. Erdman, L. M. Burke, Med. Sci. Sports Exerc. 48, 543–568 (2016).
- J. A. Hawley, C. Lundby, J. D. Cotter, L. M. Burke, Cell Metab. 27, 962–976 (2018).
- B. Egan, J. A. Hawley, J. R. Zierath, Cell Metab. 24, 342–342.e1 (2016).
- J. Bergström, E. Hultman, Nature 210, 309–310 (1966).
- L. M. Burke, L. J. C. van Loon, J. A. Hawley, J. Appl. Physiol. 122, 1055–1067 (2017).
- L. M. Burke et al., Int. J. Sport Nutr. Exerc. Metab. 28, 451–463 (2018).
- A. N. Bosch, S. C. Dennis, T. D. Noakes, J. Appl. Physiol. 76, 2364–2372 (1994).
- E. F. Coyle, A. R. Coggan, M. K. Hemmert, J. L. Ivy, J. Appl. Physiol. 61, 165–172 (1986).
- T. Stellingwerff, G. R. Cox, Appl. Physiol. Nutr. Metab. 39, 998–1011 (2014).
- J. A. Hawley, A. N. Bosch, S. M. Weltan, S. C. Dennis, T. D. Noakes, Pfluegers Arch. 426, 378–386 (1994).
- A. E. Jeukendrup, Sports Med. 47 (suppl. 1), 101–110 (2017).
- R. J. S. Costa et al., Appl. Physiol. Nutr. Metab. 42, 547–557 (2017).
- A. E. Jeukendrup, Curr. Opin. Clin. Nutr. Metab. Care 13, 452–457 (2010).
- L. M. Burke, J. A. Hawley, Med. Sci. Sports Exerc. 34, 1492–1498 (2002).
- L. M. Burke et al., Med. Sci. Sports Exerc. 34, 83–91 (2002).
- L. M. Burke, Sports Med. 45 (suppl. 1), 33–49 (2015).
- T. Stellingwerff et al., Am. J. Physiol. Endocrinol. Metab. 290, E380–E388 (2006).
- E. Maunder, A. E. Kilding, D. J. Plews, Sports Med. 48, 2219–2226 (2018).
- L. Havemann et al., J. Appl. Physiol. 100, 194–202 (2006).
- J. S. Volek et al., Metabolism 65, 100–110 (2016).
- L. M. Burke et al., J. Physiol. (London) 595, 2785–2807 (2017).
- S. D. Phinney, B. R. Bistrian, W. J. Evans, E. Gervino, G. L. Blackburn, Metabolism 32, 769–776 (1983).
- J. A. Hawley, J. J. Leckey, Sports Med. 45, S5–S12 (2015).
- J. A. Hawley, M. Hargreaves, M. J. Joyner, J. R. Zierath, Cell 159, 738–749 (2014).
- V. G. Coffey, J. A. Hawley, Sports Med. 37, 737–763 (2007).
- C. G. Perry et al., J. Physiol. (London) 588, 4795–4810 (2010).
- A. E. Jeukendrup, Sports Med. 47 (suppl. 1), 51–63 (2017).
- S. G. Impey et al., Sports Med. 48, 1031–1048 (2018).
- J. D. Bartlett, J. A. Hawley, J. P. Morton, Eur. J. Sport Sci. 15, 3–12 (2015).
- L. M. Burke, G. R. Cox, N. K. Culmmings, B. Desbrow, Sports Med. 31, 267–299 (2001).
- T. Stokes, A. J. Hector, R. W. Morton, C. McGlory, S. M. Phillips, Nutrients 10, 180 (2018).
- D. R. Moore, D. M. Camera, J. L. Areta, J. A. Hawley, Appl. Physiol. Nutr. Metab. 39, 987–997 (2014).
- P. T. Res et al., Med. Sci. Sports Exerc. 44, 1560–1569 (2012).
- G. Clénin et al., Swiss Med. Wkly. 145, w14196 (2015).
- D. J. Owens, R. Allison, G. L. Close, Sports Med. 48 (suppl. 1), 3–16 (2018).
- M. Mountjoy et al., Int. J. Sport Nutr. Exerc. Metab. 28, 316–331 (2018).
- L. M. Burke, B. Lundy, I. L. Fahrenholtz, A. K. Melin, Int. J. Sport Nutr. Exerc. Metab. 28, 350–363 (2018).
- T. Stellingwerff, Int. J. Sport Nutr. Exerc. Metab. 28, 428–433 (2018).
- S. Bermon et al., Exerc. Immunol. Rev. 23, 8–50 (2017).
- C. Sale et al., J. Appl. Physiol. 119, 824–830 (2015).
- A. K. A. McKay et al., Med. Sci. Sports Exerc. 10.1249/ MSS.0000000000001816 (2018).
- T. L. Merry, M. Ristow, J. Physiol. (London) 594, 5135–5147 (2016).
- W. K. Yeo et al., J. Appl. Physiol. 105, 1462–1470 (2008).
- S. C. Lane et al., J. Appl. Physiol. 119, 643–655 (2015).
- L. A. Marquet et al., Med. Sci. Sports Exerc. 48, 663–672 (2016).
- K. D. Gejl et al., Med. Sci. Sports Exerc. 49, 2486–2497 (2017).
- T. Stellingwerf, Int. J. Sport Nutr. Exerc. Metab. 22, 392–400 (2012).
- D. Bishop, Sports Med. 40, 995–1017 (2010).
- G. Cox, I. Mujika, D. Tumilty, L. Burke, Int. J. Sport Nutr. Exerc. Metab. 12, 33–46 (2002).
- J. A. Hawley, E. J. Schabort, T. D. Noakes, S. C. Dennis, Sports Med. 24, 73–81 (1997).
- E. D. B. Goulet, M. Aubertin-Leheudre, G. E. Plante, I. J. Dionne, Int. J. Sport Nutr. Exerc. Metab. 17, 391–410 (2007).
- R. W. Kenefick, Sports Med. 48 (suppl. 1), 31–37 (2018).
- M. Z. Donahue, “Runner comes excruciatingly close to breaking two-hour marathon banner,” 6 May 2017; https://news.nationalgeographic.com/2017/05/extreme-running-marathonnike-science/.
- M. J. Joyner, J. R. Ruiz, A. Lucia, J. Appl. Physiol. 110, 275–277 (2011).
- M. J. Joyner, E. F. Coyle, J. Physiol. (London) 586, 35–44 (2008).
- W. Hoogkamer et al., Sports Med. 48, 1009–1019 (2018).
- A. M. Jones, Appl. Physiol. Nutr. Metab. 39, 1019–1028 (2014).
- A. Krogh, J. Lindhard, Biochem. J. 14, 290–363 (1920).
- L. M. Burke, P. Peeling, Int. J. Sport Nutr. Exerc. Metab. 28, 159–169 (2018).
- R. K. Boorsma, J. Whitfield, L. L. Spriet, Med. Sci. Sports Exerc. 46, 2326–2334 (2014).
- K. L. Jonvik, J. Nyakayiru, L. J. van Loon, L. B. Verdijk, J. Appl. Physiol. 119, 759–761 (2015).
- M. Hultström et al., J. Appl. Physiol. 119, 762–769 (2015).
- L. Y. Beis et al., J. Int. Soc. Sports Nutr. 8, 7 (2011).
- V. O. Onywera, F. K. Kiplamai, P. J. Tuitoek, M. K. Boit, Y. P. Pitsiladis, Int. J. Sport Nutr. Exerc. Metab. 14, 709–719 (2004).
- J. P. Morton, C. Robertson, L. Sutton, D. P. MacLaren, Int. J. Sport Nutr. Exerc. Metab. 20, 80–85 (2010).
- T. Fordyce, “Chris Froome: Team Sky’s unprecedented release of data reveals how British rider won Giro d’Italia,” 4 July 2018; https://www.bbc.com/sport/cycling/44694122.
- L. Y. Beis, M. Wright-Whyte, B. Fudge, T. Noakes, Y. P. Pitsiladis, Clin. J. Sport Med. 22, 254–261 (2012).
- G. R. Cox et al., J. Appl. Physiol. 93, 990–999 (2002).
- A. Jeukendrup, Sports Med. 44 (suppl. 1), S25–S33 (2014).
- F. A. Bainbridge, The Physiology of Muscular Exercise (Longmans, Green & Co., 1919).
- J. A. Hawley, R. J. Maughan, M. Hargreaves, Cell Metab. 22, 12–17 (2015).
- S. A. Levine, B. Gordon, C. L. Derick, JAMA 82, 1778–1779 (1924).
- T. Matsui et al., J. Physiol. (London) 590, 607–616 (2012).
- L. L. Spriet, Sports Med. 44 (suppl. 2), S175–S184 (2014).
- L. M. Burke, R. J. Maughan, Eur. J. Sport Sci. 15, 29–40 (2015).
- J. M. Carter, A. E. Jeukendrup, D. A. Jones, Med. Sci. Sports Exerc. 36, 2107–2111 (2004).
- E. S. Chambers, M. W. Bridge, D. A. Jones, J. Physiol. (London) 587, 1779–1794 (2009).
- S. Gam, K. J. Guelfi, P. A. Fournier, Sports Med. 46, 1385–1390 (2016).
- D. H. Craighead et al., Muscle Nerve 56, 379–385 (2017).
- Persistence Market Research, “Global market study on sports supplements: Non-protein products to witness substantial growth during 2017 – 2025” (Rep. PMRREP3034, Persistence Market Research, January 2018); https://www.persistencemarketresearch.com/market-research/sportssupplements-market.asp.
- I. Garthe, R. J. Maughan, Int. J. Sport Nutr. Exerc. Metab. 28, 126–138 (2018).
- R. J. Maughan et al., Int. J. Sport Nutr. Exerc. Metab. 28, 104–125 (2018).
- P. Peeling, M. J. Binnie, P. S. R. Goods, M. Sim, L. M. Burke, Int. J. Sport Nutr. Exerc. Metab. 28, 178–187 (2018).
- A. I. Geller et al., N. Engl. J. Med. 373, 1531–1540 (2015).
- World Anti-Doping Agency, The Code; https://www.wada-ama.org/en/what-we-do/the-code.
- J. M. Martínez-Sanz et al., Nutrients 9, 1093 (2017).
- P. R. Below, R. Mora-Rodríguez, J. González-Alonso, E. F. Coyle, Med. Sci. Sports Exerc. 27, 200–210 (1995).
- S. A. Conger, G. L. Warren, M. A. Hardy, M. L. Millard-Stafford, Int. J. Sport Nutr. Exerc. Metab. 21, 71–84 (2011).
- L. M. Burke, Sports Med. 47 (suppl. 1), 79–100 (2017).
- A. K. Garth, L. M. Burke, Sports Med. 43, 539–564 (2013).
- R. Reale, G. Slater, L. M. Burke, Int. J. Sports Physiol. Perform. 13, 459–466 (2018).
- C. Heneghan, R. Perera, D. Nunan, K. Mahtani, P. Gill, BMJ 345, e4797 (2012).
- C. Heneghan et al., BMJ 345, e4848 (2012).
- M. N. Sawka et al., Med. Sci. Sports Exerc. 39, 377–390 (2007).
- D. Cohen, BMJ 345, e4737 (2012).