

17 Jul El máximo estado estable del lactato: Redefinición de la metodología de referencia (gold standard)
Andrew M. Jones1, Mark Burnley2, Matthew I. Black1, David C. Poole3y Anni Vanhatalo1
1 Sport and Health Sciences, University of Exeter, St. Luke’s Campus, Exeter, United Kingdom
2 School of Sport and Exercise Sciences, University of Kent, Medway, United Kingdom
3 Department of Kinesiology, Kansas State University, Manhattan, Kansas
Resumen
El máximo estado estable del lactato (MLSS) y la potencia crítica (CP) son dos índices muy utilizados de la tasa metabólica oxidativa más alta que se puede mantener durante un ejercicio continuo y, a menudo, se considera que son sinónimos.Sin embargo, a pesar de que poseen similitudes en sus fundamentos, las diferencias metodológicas que existen en la valoración de estos parámetros generalmente hacen que el MLSS se alcance en una producción de potencia o velocidad de carrera algo mas bajas y que un ejercicio en laCP sea sostenible durante no más de aproximadamente 20-30 minutos. Esto ha llevado a la opinión de que la CP sobreestima el máximo estado estable metabólico “real” y que el MLSS debe ser considerado la metodología de referencia o “gold standard” para la evaluación de la capacidad de realizar ejercicios de resistencia.En este artículo presentaremos evidencia consistente con la conclusión contraria: esto es que (1) tal como se lo define en la actualidad, el MLSS subestima naturalmente el máximo estado estable metabólico real;y (2) la CP solo representa el límite entre dominios de intensidad de ejercicio discretos dentro de los cuales las respuestas metabólicas dinámicas cardiorrespiratorias y musculares que se producen frente al ejercicio difieren profundamente.Si bien el MLSS y la CP pueden tener relevancia para el entrenamiento y el rendimiento deportivo, buscamos que se aprecie mejor la distinción entre los dos conceptos/métricas, y que se detengan las comparaciones entre MLSS y CP surgidas de la creencia errónea de que son teóricamente sinónimos.La CP representa el ejercicio que permite la delimitación de límites genuinos entre ejercicios donde se puede mantener la homeostasis fisiológica y ejercicios en los que no se puede, y debe ser considerado la metodología de referencia o “gold standard” cuando el objetivo es determinar el máximo estado estable metabólico.
Palabras clave: Fatiga, metabolismo, rendimiento
Introducción
La determinación de la velocidad de carrera o producción de potencia de ciclismo que genera la máxima tasa metabólica oxidativa sostenible puede ser importante para evaluar el potencial de rendimiento deportivo y para guiar los programas de entrenamiento deportivo (Jones y Cárter 2000; Morton 2006; Jones et al. 2010; Vanhatalo et al. 2011a).Al realizar entrenamiento por debajo de este umbral en comparación con entrenamiento por encima del mismo, se producirándiferencias agudas en el suministro de energía oxidativa y no oxidativa, en la bioquímica muscular y sanguínea, en las respuestas cardiorrespiratorias, en los procesos de fatiga y en la percepción del esfuerzo, que, si se repiten crónicamente, probablemente promuevan diferentes adaptaciones fisiológicas (Holloszy y Coyle 1984; Jones y Cárter 2000).Si bien esta noción es ampliamente aceptada, han surgido una gran cantidad de términos y técnicas que pretenden describir o determinar este “máximo estado estable metabólico”.Por ejemplo,entre los fenómenos derivados de los tests de ejercicio incremental, que se proponen como equivalentes a, o que permiten una estimación precisa del, máximo estado estable metabólico se incluyen el umbral de lactato (LT), el umbral de intercambio de gases (GET), el umbral ventilatorio, el punto de inflexión del lactato ( LTP), el umbral anaeróbico, el “inicio de la acumulación de lactato en sangre” que coincide con una concentración absoluta de lactato en sangre de 4 mmol/L (OBLA), umbral anaeróbico individual, mínimo de lactato y umbral de compensación respiratoria (Faude et al. 2009; Jones et al. 2018).Estos términos no solo reflejan eventos y mecanismos fisiológicos muy diferentes, sino que pueden producirse en tasas metabólicas diferentes y, en cierta medida, pueden depender de la técnica de medición o del paradigma de prueba específico que se haya utilizado (Jamnick et al. 2018).En consecuencia, esto ha provocado una considerable confusión y malentendido en este campo (ver Jones et al. 2018, para obtener mas detalles).
Como primer paso para contar con un panorama mas claro, es esencial apreciar la existencia de dominios discretos de intensidad de ejercicio dentro de los cuales las respuestas fisiológicas al ejercicio difieren considerablemente (Whipp y Ward 1992; Poole y Richardson 1997; Cárter et al. 2002; Wilkerson et al. 2004; Black et al. 2017).Las respuestas del consumo de oxígeno pulmonar (VO2) y de la concentración de lactato sanguíneo frente a ejercicios de potencia constante e intensidad moderada, alta y severa se presentan en la Figura 1. Estos perfiles indican que el arribo a valores de estado estable en estas variables es marcadamente diferente en estos dominios de ejercicio discreto (i.e. rápida en el dominio de intensidad moderada, retrasada en el dominio de intensidad alta y no es posible en el dominio de intensidad severa).Estas diferencias reflejan la contribución variable de los sistemas de energía y tienen claras implicaciones para el desarrollo de la fatiga y la tolerancia al ejercicio (Whipp y Ward 1992; Jones et al. 2011).Algunos “umbrales” identificables durante el ejercicio incremental marcan la transición de ejercicio de intensidad moderada a intensa (i.e., LT y GET), mientras que otros delimitarían la transición del ejercicio de intensidad alta a severa (i.e., LTP y, posiblemente, OBLA). El primer umbral es relevante para eventos de ultra resistencia y eventos de resistencia de baja intensidad y en fisiología ocupacional y clínica.Sin embargo, el objeto de este trabajo de revisión es la definición, la evaluación y la aplicación del segundo límite, que generalmente se produce en el 75-90% del VO2max y, por lo tanto, es más importante para el rendimiento de la mayoría de los tipos de ejercicios de resistencia de alto nivel (Jones y Poole 2008; Jones y Vanhatalo 2017).
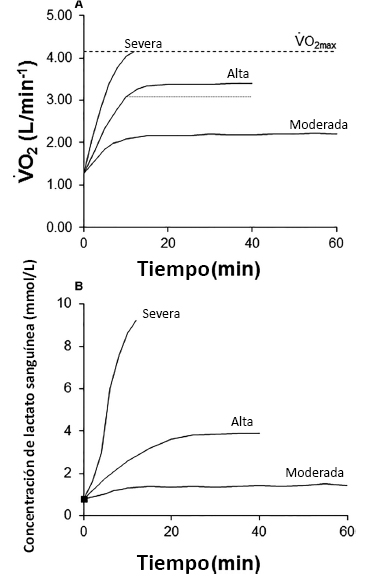
Figura 1. Representación esquemática de las respuestas del consumo de oxígeno pulmonar (VO2)(Panel A) y de la concentración de lactato sanguíneo (Panel B) durante el ejercicio de intensidad moderada, alta y severa.Durante el ejercicio de intensidad moderada, el VO2 y la concentración sanguínea de lactato alcanzan valores de estado estable de forma brusca.Durante el ejercicio de alta intensidad y el de intensidad severa, hay un costo adicional de oxígeno (denominado componente lento del VO2) superior a lo esperado a partir de la extrapolación de la relación entre el VO2 de intensidad moderada y la producción de potencia.Durante el ejercicio de alta intensidad, se retrasa el momento en que se alcanzan (mayor amplitud) los valores de estado estable para el VO2 y la concentración de lactato sanguínea.La magnitud del componente lento del VO2 durante el ejercicio de alta intensidad se puede observar en el panel A representado con la línea punteada. Durante el ejercicio de intensidad severa, el VO2 y la concentración de lactato sanguíneo continúan incrementándose hasta que se alcanza el VO2máx (Panel A, línea discontinua) y el límite de tolerancia se produce poco tiempo después.
La metodología adecuada para la determinación del máximo estado estable metabólico (es decir, la velocidad umbral o producción de potencia que separa un ejercicio de alta intensidad de uno de intensidad severa) es controversial. Frecuentemente se considera que el “gold standar” es el denominado máximo estado estable del lactato(MLSS; Beneke y von Duvillard 1996; Billat et al. 2003; Faude et al. 2009). El MLSS se obtiene convencionalmentea partir deuna serie (típicamente 4-5) de ejercicios continuos de 30 min, realizadas en días separados, con velocidades de carrera o producciones de potencia diferentes pero constantes;la concentración de lactato sanguíneo se mide en reposo y después de cada 5 minutos de ejercicio y el MLSS se define como el valor mas alto de velocidad o de producción de potencia que no produce un aumento en la concentración de lactato en sangre superior a 1mmol/L entre 10 y 30 min (Beneke 1995; Jones y Doust 1998; ver Figura 2).Alternativamente, el máximo estado estable metabólico puede ser determinado mediante la potencia crítica (CP o velocidad crítica durante la carrera1), que se obtiene a partir de la relación hiperbólica entre la velocidad o la producción de potencia y la duración durante la cual se puede mantener esa velocidad o producción de potencia (Hill 1925; Monod y Scherrer 1965; Hill y Smith 1999; Hill et al. 2002; Jones et al. 2010; ver la Figura 3).
1Notamos aquí que el término “intensidad” (por ejemplo, “intensidad crítica”) es inapropiado en este contexto.Esto se debe a que a una velocidad o producción de potencia constante dada, la intensidad del ejercicio (es decir, la fracción de VO2max requerida o la perturbación metabólica muscular provocada) puede cambiar; esto es especialmente cierto para el ejercicio cerca de la velocidad / potencia crítica donde se espera un comportamiento en estado no estacionario.También se sabe que la producción de potencia (la tasa de transferencia de energía desde el músculo esquelético para realizar un trabajo externo) y la intensidad del ejercicio (la magnitud de la fluctuación (s) metabólica (s) provocada por la tarea) se pueden disociar completamente dependiendo de la duración trabajo:recuperación durante el ejercicio intermitente, en comparación con el ejercicio continuo (Davies et al. 2017).Por lo tanto, es preferible utilizar el término ‘crítico’ junto a la unidad SI asociada que sea apropiada para la modalidad de ejercicio (potencia, velocidad o velocidad, torque etc.).
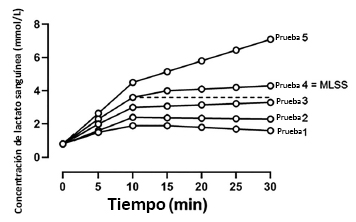
Figura 2. Representación esquemática de la respuesta de la concentración de lactato sanguíneo frente a una serie de pruebas de velocidad de carrera constante realizadas en días separados para la determinación de MLSS. La Prueba 1 es representativa de la velocidad de carrera más baja elegida y cada prueba es indicativa de un incremento en la velocidad hasta la Prueba 5 (la velocidad de carrera más alta aplicada).Durante las Pruebas 1, 2, 3 y 4, la concentración sanguínea de lactato no aumenta más de 1 mmol/L entre los minutos 10 y 30. Sin embargo, durante la prueba 5, la concentración de lactato sanguíneo es de 4,5 mmol/L a los 10 minutos y 7,1 mmol/L a los 30 min (∆2,6 mmol/L).Por lo tanto, a pesar de un aumento gradual (∆0,7 mM) en la concentración de lactato sanguíneo entre los minutos 10 y 30, la prueba 4 representa la mayor velocidad de carrera en la cual la concentración del lactato en sangre no aumenta mas de 1 mM, y por lo tanto este podría ser definido como MLSS.Tenga en cuenta, por lo tanto, que el MLSS real, según la definición aceptada, se encuentra a una velocidad en algún lugar entre la prueba 4 y la prueba 5, de modo que el MLSS seleccionado (Prueba 4) será necesariamente una subestimación.La línea discontinua es indicativa de la concentración de lactato en sangre obtenida a los 10 minutos durante la Prueba 4, y se proyecta hasta el final del ejercicio.
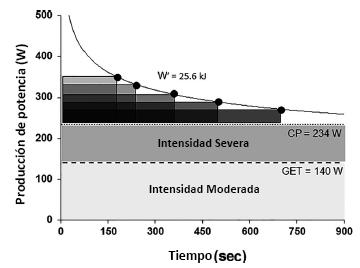
Figura 3. Representación esquemática de la relación potencia-duración con referencia a los dominios de intensidad de ejercicio de intensidad moderada (área sombreada con gris claro) y de alta intensidad (área sombreada con gris oscuro).El límite entre los dominios de intensidad moderada y alta está dado por el umbral del lactato o umbral de intercambio de gases (GET), y el límite entre los dominios de intensidad alta y severa está dado por la potencia crítica (CP). La CP y la capacidad de trabajo disponible por encima de la misma (denominado W´) pueden ser determinadas utilizando una serie de pruebas de potencia constante realizadas hasta el límite de tolerancia dentro del dominio de intensidad severa (es decir,> CP). La CP se define como la asíntota de potencia (234 W en este ejemplo), y W´ se caracteriza por la constante de la curva (25,6 kJ en este ejemplo), de esta relación hiperbólica entre la producción de potencia y el tiempo.La W´ es la capacidad, pero no la tasa, es limitada y, por lo tanto, su contribución (en kJ) al ejercicio de intensidad severa es constante independientemente de la duración del ejercicio en el dominio de intensidad severa.Cuanto mayor sea la diferencia entre la producción de potencia sostenida y la CP, más rápidamente se utilizará W´, y el límite de tolerancia coincidirá con el agotamiento de W´. La relación hiperbólica entre potencia y tiempo puede transformarse en relación lineal dibujando el trabajo realizado en función del tiempo, en cuyo caso la pendiente de la línea representa la CP y la ordenada al origen representa W´, o la potencia en función 1/tiempo, en cuyo caso la pendiente de la línea representará W´ y la ordenada al origen será la CP.
El propósito de este artículo es realizar un análisis crítico de los conceptos de MLSS y CP y evaluar su validez en la determinación del máximo estado estable metabólico. Aunque el MLSS y la CP tienen algunas similitudes conceptuales, tienden a aproximarse entre sí y frecuentemente se afirma que representan el mismo fenómeno, en la práctica las diferencias metodológicas en su determinación suelen producir divergencias (es decir, MLSS <CP).Por lo tanto, es importante destacar que apreciar las posibles ventajas y desventajas de, y las posibles diferencias y similitudes entre MLSS y CP, no es simplemente una cuestión de semántica.Por el contrario, este problema es sumamente importante porque tiene el potencial de influir en el pronóstico de rendimiento y en la prescripción de ejercicios/entrenamiento. Por lo tanto, es relevante no solo para los investigadores deportivos y para las ciencias deportivas sino que también paraatletas, entrenadores y profesionales del ejercicio.Sostenemos que: (1) MLSS y CP ya no deben ser considerados como sinónimos; y (2) la CP tiene los fundamentos teóricos y la base de evidencia más sólidos, y en lo sucesivo debe ser considerado la metodología de referencia o “gold standard” para definir el máximo estado estable metabólico, es decir, el límite entre los dominios de ejercicio de intensidad alta y severa.
Consideraciones sobre la Definición y Determinación del MLSS
El origen del concepto MLSS es un tanto oscuro, pero tal vez se pueda atribuir al trabajo de los fisiólogos alemanes, Mader y Heck, en la década de 1980 (Heck et al. 1985; Mader and Heck 1986).Inicialmente, se consideró que el MLSS se producía en una concentración de lactato sanguíneo igual a 2,2 mmol/L (LaFontaine et al. 1981; Priest y Hagan 1987) o, más a menudo, 4 mmol/L (Sjódin et al. 1982; Stegmann y Kin dermann 1982; Heck et al. 1985; Mader y Heck 1986).Sin embargo, los descubrimientos de que la concentración de lactato sanguíneo absoluta en MLSS variaba considerablemente entre individuos (Beneke y von Duvillard 1996) y entre las modalidades de ejercicio (Beneke et al. 2001) hicieron que posteriormente el MLSS sea reconsiderado para representar la velocidad o producción de potencia en MLSS, independientemente de la concentración de lactato sanguíneo absoluta.Yamamoto et al.(1991) definieron el MLSS como la mayor producción de potencia en la cual la concentración de lactato en sangre no aumentaba luego de 15 a 30 min de ejercicio continuo y señalaron que “a pesar de la arbitrariedad de la definición, el MLSS podría ser útil para prescribir ejercicio prolongado porque se puede hacer ejercicio sin la acumulación continua de lactato en sangre durante al menos 30 minutos “.Estudios posteriores introdujeron una definición modificada de MLSS, que sigue siendo ampliamente utilizada: la producción de potencia más alta en la cual el aumento en la concentración de lactato sanguíneo es inferior a 1 mmol/L después de entre 10 y 30 minutos de ejercicio (Snyder et al. 1994; Beneke y von Duvillard 1996; Jones y Doust 1998; Beneke et al. 2000). Beneke (2003) informó que la determinación de MLSS dependería del protocolo, donde las sesiones de ejercicios de 30 minutos producirían una producción de potencia más baja en el MLSS que las sesiones de ejercicios de 20 minutos.Sin embargo, no está clara la razón para la definición altamente específica, pero aparentemente arbitraria, de MLSS, que contempla el período de 10-30 minutos y la magnitud aceptable del cambio en la concentración de lactato sanguíneo.
Hay varias preocupaciones metodológicas con respecto a la evaluación de MLSS que es necesario destacar.Que una velocidad o producción de potencia en particular se encuentre por encima o por debajo del MLSS depende esencialmente de solo dos mediciones de la concentración del lactato en sangre, generalmente a partir de una muestra de sangre que se obtiene de la yema de los dedos o del lóbulo de la oreja, una obtenida a los 10 minutos y la otra a los 30 minutos de ejercicio.Si el aumento en la concentración de lactato sanguínea es <1 mmol/L, entonces la velocidad o la producción de potencia se consideraque está por debajo de MLSS, mientras que si el aumento es > 1 mmol/L, entonces se considera que la velocidad o la producción de potencia están por encima del MLSS.Sin embargo, es necesario tener en cuenta que la medición de la concentración del lactato en sangre por medio de analizadores de uso general generalmente tiene un error de 0,2-0,4 mmol/L (Bonaventura et al. 2015) y que la fiabilidad de la medición de la concentración de lactato sanguíneo, que representa una combinación de variación biológica y error analítico, durante la prueba de ejercicio submáxima es del 11-52% (Saunders et al. 2004).Con tal inexactitud potencial, es obvio que el potencial de obtener falsos positivos, es decir, que la velocidad o la potencia se consideren por encima de MLSS cuando no lo están, o falsos negativos, es decir, que se considere que la velocidad o la potencia están por debajo de MLSS cuando no lo están, es bastante alto. También se debe tener en cuenta que el MLSS se ve afectado por breves interrupciones en el ejercicio que a menudo son necesarias para facilitar el muestreo de sangre (Beneke et al. 2003).Además, 30 minutos de ejercicio de intensidad alta a severa pueden hacer que la sangre se concentre como consecuencia de la variación en el nivel de fluidos y por la deshidratación relacionada con el sudor que, si no se corrige, tendrá un impacto adicional en la concentración del lactato que se determine, al menos si se mide en sangre completa (Dill y Costill 1974).Una complicación adicional cuando se prolonga la duración del ejercicio para una determinada velocidad o producción potencia dentro de este dominio de intensidad es el conocido cambio en la utilización del sustrato desde carbohidratos hacia el metabolismo de los ácidos grasos (Hermansen et al. 1967), una adaptación que tenderá a reducir la producción de lactato muscular.Es importante destacar que no es seguro que la concentración de lactato en sangre en un instante dado refleje adecuadamente el estado metabólico del músculo que está realizando ejercicio (Jorfeldt et al. 1978; Tesch et al. 1982; Bergman et al. 1999).La interacción dinámica entre las tasas de producción de lactato muscular, el flujo del lactato desde el músculo hacia la sangre y el aclaramiento/metabolismo del lactato tanto dentro del músculo como en la sangre por parte de otros órganos (Stainsby y Brooks 1990), significa que un estado estable en la concentración sanguínea de lactato no necesariamente implica la existencia de un estado estable bioenergético en el músculo esquelético que se está contrayendo.También existe evidencia de que la dinámica de la concentración del lactato sanguíneo se puede disociar de la tasa metabólica oxidativa del cuerpo entero: hay ejemplos de perfiles con concentraciones de lactato sanguíneo elevada y crecientes en presencia de un VO2 claramente en estado estable (Scheen et al. 1981);y se ha demostrado que la infusión de epinefrina altera la dinámica de la concentración del lactato en sangre sin producir cambios en el VO2(Gaesser et al. 1994; Womack et al. 1995).En conjunto, estos puntos sugieren que la concentración del lactato sanguíneo, per se,no es ni una medida adecuada ni lo suficientemente sensible para permitir una evaluación confiable en referencia a si una velocidad o producción de potencia específicas pueden ser sostenibles en un estado metabólico estable.
Otros aspectos del protocolo de evaluación del MLSS también deben ser discutidos.Durante el ejercicio sostenido de alta intensidad, la concentración de lactato sanguíneo tiende a aumentar de forma curvilínea con el tiempo, y la tasa de cambio de la concentración de lactato sanguíneo tiende a aumentarmás en los primeros 5-10 minutos que en los últimos 5-10 minutos de una sesión de ejercicios de 30 minutos (Figura 4; Scheen et al. 1981; Jones y Doust 1998).Esto tiene el potencial de producir un escenario en el que se considera que la velocidad o la producción de potencia están por encima de MLSS a pesar de que la concentración de lactato sanguíneo se mantiene estable (o incluso disminuye) durante los últimos 10-15 minutos del ejercicio de 30 minutos, y un perfil de este tipo debería interpretarse como un indicador de que se alcanzó un estado estable retrasado.La evaluación de MLSS también depende de sujetos que realizan una serie de ejercicios en diferentes días a velocidades o producciones de potencia discretas que generalmente difieren en 1 km/h o 10-30 W, respectivamente (Jones y Doust 1998; Cárter et al. 1999; Smith y Jones 2001; Pringle y Jones 2002; Beneke 2003; Iannetta et al. 2018).Por definición, la selección de MLSS debe realizarse a una de estas velocidades o producciones de potencia discretas, con el resultado inevitable de que el MLSS seleccionado siempre debe ser más bajo que el MLSS real.Por ejemplo, si el comportamiento de la concentración de lactato sanguíneo indica que la velocidad de 16 km/h está por debajo del MLSS y la velocidad de 17 km/h por encima del MLSS, entonces se seleccionará 16 km/h para el MLSS.Sin embargo, si se hubiera aplicado, una velocidad de 16,5 km/h también podría haber producido una respuesta de concentración de lactato sanguíneo consistente con el ejercicio por debajo del MLSS, de modo que 16,5 km/h hubiera sido seleccionado como MLSS.En término medio, con diferencias de 1 km/h o 30 W entre pruebas discretas, el MLSS será subestimado en 0,5 km/h en runningo en 15 W en ciclismo, respectivamente.Por lo tanto lalimitadagranularidadinherente en el protocolo MLSS, inevitablemente produce una subestimación del MLSS “real”.De hecho, es crucial apreciar que, tal como se define y se mide actualmente, el MLSS deberesidirdentro del dominio de intensidad alta y no en el límitede los dominios de intensidad alta y severa.
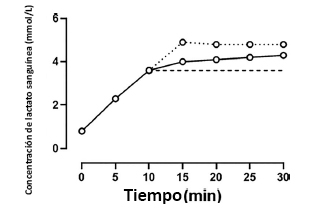
Figura 4. Respuesta de la concentración de lactato sanguíneo en una prueba de producción de potencia constante indicativa del MLSS (línea negra continua) frente a una respuesta de la concentración de lactato sanguíneo que sería (de acuerdo con la definición estricta de MLSS que considera sólo los valores absolutos de concentración del lactato en sangre a los 10 y 30 minutos), establecido por encima de MLSS (línea punteada). Sin embargo, debemos tener en cuenta que a pesar de ubicarse supuestamente por encima de MLSS (línea punteada), la concentración de lactato sanguíneo se estabilizó entre los 15 y 30 min.Esta es una de las fuentes potenciales de error en la definición del MLSS a partir de solo dos datos puntuales y de la aplicación de un limite de tolerancia arbitrario (∆1,0 mmol/L) para el aumento de concentración de lactato sanguíneo entre estos dos puntos.
La precisión de la estimación del MLSS se incrementa naturalmente al usar diferencias de producciones de potencia o de velocidad más pequeñas entre una prueba y la siguiente (por ejemplo, 0,5 km/h para running, 15 W para ciclismo), pero es probable que esta metodología incremente el número de pruebas necesarias para la determinación del MLSS.Debido a que la concentración de lactato sanguíneo es sensible a las variaciones en la hidratación y en el estado nutricional del individuo (Jacobs 1986), particularmente en términos de niveles de glucógeno muscular, los sujetos deben abstenerse de entrenar normalmente y deben consumir una dieta consistente durante el período de prueba, que podría extenderse más de cinco días.Esto limita la practicidad de evaluar el MLSS con fines de investigación y en el trabajo aplicado con atletas.Además, al menos en los sujetos menos entrenados, el extenso protocolo requerido para la evaluación del MLSS podría ser lo suficientemente arduo como para estimular las adaptaciones al entrenamiento que producen un aumento de MLSS.Se han propuesto enfoques que pretenden permitir una evaluación MLSS precisa a partir de una menor cantidad de pruebas (por ejemplo, Billat et al., 1994; Kilding y Jones, 2005), pero estos no estarían libres de las críticas sobre la evaluación del MLSS que efectuamos más arriba.
Consideración de la Potencia Crítica (CP) como la Metodología de Referencia o “Gold Standard” Adecuada para Determinar el Máximo Estado Estable Metabólico
La potencia crítica (CP) tiene fundamentos históricos, teóricos, fisiológicos y matemáticos sólidos (Hill 1925; Wilkie 1960; Monod y Scherrer 1965; Morilani el al. 1981; Poole el al. 1988; Hill el al. 2002; Morton 2006; Jones et al. 2008; Vanhatalo et al. 2016; Mitchell et al. 2018).De hecho, la hiperbolicidad de la relación entre la velocidad o la producción de potencia y la duración durante la cual es posible mantener la velocidad o la producción de potencia fue reconocida por primera vez por AV Hill en 1925, mientras realizaba un seguimiento de los rendimientos de records mundiales de diferentes deportes (Hill 1925; para obtener mas datos ver Figura 5).En la actualidad esta función hiperbólica, con su asíntota y constante de curvatura inherentes, es reconocida como una propiedad bioenergética de los sistemas vivientes, y ha sido descrita en múltiples especies (Full y Herreid 1983; Full 1986; Lauderdale y Hinchcliff 1999; Billat et al .2005; Copp et al. 2010) y en modalidades de ejercicio de músculos aislados y de todo el entero (Monod y Scherrer 1965; Hughson el al. 1984; Poole el al. 1988; Burnley 2009).
La CP es única en lo que respecta a los “umbrales” fisiológicos, ya que, si bien representa una tasa metabólica critica (Barker el al. 2006; Vanhatalo et al. 2016), su definición se basa únicamente en la medición del trabajo mecánico realizado y en la tolerancia al ejercicio. Sin embargo lo más importante es que la PC separa dos dominios de ejercicio que se caracterizan por un comportamiento fisiológico distinto.Quizás lo más importante es que la CP representa un límite por encima del cual el ejercicio permite alcanzar el VO2máx, siempre que el ejercicio pueda mantenerse durante el tiempo suficiente (es decir, con una duración igual o mayor a aproximadamente 2 min) para que se pueda alcanzar este valor (Poole et al. 1988; Hill y Ferguson 1999; Hill y Smith 1999; Hill et al. 2002; Vanhatalo et al. 2016).La magnitud de la diferencia entre la producción de potencia que se puede mantener y la CP determinará la tasa la cual se utilizará la capacidad de trabajo limitada por encima de CP (W´)pero, para cualquier sesión de ejercicio en el dominio de intensidad severa, el límite de tolerancia coincidirá con el agotamiento de W´ y el alcance simultáneo del VO2max(Murgatroyd et al. 2011; Vanhatalo et al. 2011b).Esto significa que el tiempo hasta el límite de tolerancia para cualquier producción de potencia en el dominio de intensidad severa se puede calcular con precisión solamente conociendo la producción de potencia que se mantendrá y la CP y W´ del individuo (Vanhatalo et al. 2011a; Jones and Vanhatalo 2017). Además a diferencia del MLSS, en el cual el cambio en la concentración de lactato sanguíneo es el único índice según el cual se clasifica el comportamiento fisiológico no estacionario, se ha demostrado que la CP separa los dominios discretos de intensidad de ejercicio que tienen diferentes perfiles de metabolismo muscular (Jones et al. 2008; Vanhatalo et al. 2016; Black et al. 2017), neuromusculares ( Burnley et al. 2012; Black et al. 2017), de intercambio de gases respiratorios y ventilación (Poole et al. 1988; Murgatroyd et al. 2014), cardiovasculares (Copp et al. 2010) y ácido-base de la sangre (incluida la concentración de lactato sanguíneo Poole et al. 1988; Pringle y Jones 2002; Vanhatalo et al.2016).Estas diferencias globales en el comportamiento fisiológico por encima y por debajo de la CP se resumen en la Figura 6.
Varios estudios compararon el MLSS y la CP determinados independientemente e informaron que la CP se produce en una producción de potencia mayor a la del MLSS (Jenkins y Quigley 1990; Smith y Jones 2001; Pringle y Jones 2002; Dekerle et al. 2003, 2005; Mattioni Maturana et al. 2016; cf. Keir et al. 2015).En promedio, se ha informado que la PC es un 7% más alta que el MLSS (p. ej., 4%, Smith y Jones 2001; 9%, Pringle y Jones 2002; 16%, Dekerle et al. 2003; 5%, Dekerle et al.2005;9%, Mattioni Maturana et al.2016;cf.1%, Keir et al.2015).Dado que el MLSS naturalmente subestimará el límite entre el ejercicio de intensidad alta y el de intensidad severa, y teniendo en cuenta la magnitud del error asociado con la determinación tanto del MLSS como de la CP, esta diferencia no debe ser considerada sorprendente.Sin embargo, estos estudios han interpretado la diferencia entre MLSS y CP como evidencia de que la CP no representa la tasa metabólica oxidativa sostenible más alta, con el supuesto inherente de que MLSS es la metodología de referencia.Tal como se mencionó previamente, una interpretación alternativa es que la falta de concordancia entre MLSS y CP indica que MLSS subestima la tasa metabólica oxidativa más alta en estado estable.De acuerdo con esto, se informó que el VO2durante el ejercicio continuo realizado a 10 W por encima de MLSS permitió alcanzar un VO2de estado estableequivalente a aprox. 90% de VO2pico(Mattioni Maturana et al. 2016; Iannetta et al. 2018), un comportamiento que claramente es indicativo de ejercicio de alta intensidad.
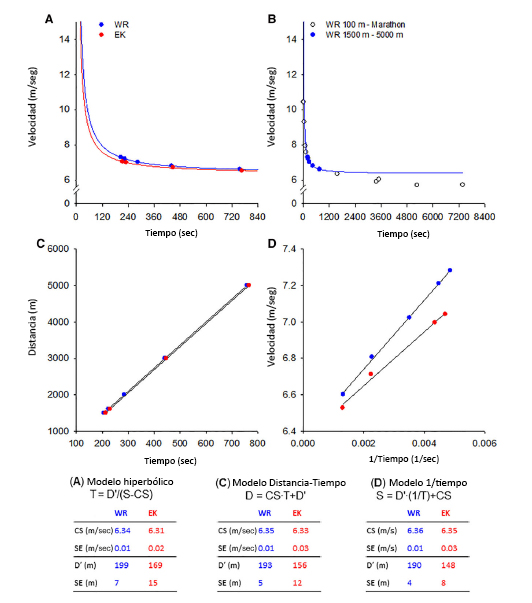
Figura 5. El Panel A presenta la relación hiperbólica entre velocidad y tiempo trazada para los récords mundiales actuales (a partir de marzo de 2019) en 1500 m hasta 5000 m (en azul se presentan los records obtenidos por diferentes atletas) y las mejores marcas personales en las mismas distancias recorridas por un corredor de distancia de élite individual (Eliud Kipchoge, EK, en rojo).El Panel B demuestra que la curva hiperbólica construida para los records mundiales en 1500 m a 5000 m (en azul, los mismos datos que en el Panel A) no se ajusta bien a los rendimientos obtenidos por los records mundiales de distancias más cortas (100 a 800 m) o más largas (10000 m hasta el maratón).Por lo tanto, la relación hiperbólica es válida para eventos cuya duración va desde ~ 2 min hasta quizás 15-20 min.La transformación lineal de la relación hiperbólica se muestra en el Panel C (gráfico distancia-tiempo donde la pendiente de la línea de regresión lineal es la velocidad crítica, CS, y la ordenada al origen equivale a la constante de la curva, D’) y el Panel D (grafica de la relación velocidad-1/ tiempo donde la pendiente representa a D’ y la ordenada al origen a CS). Al pie de la figura se presentan las estimaciones de CS y D’ obtenidas a partir de las tres ecuaciones, con los errores estándar asociados a la estimación.
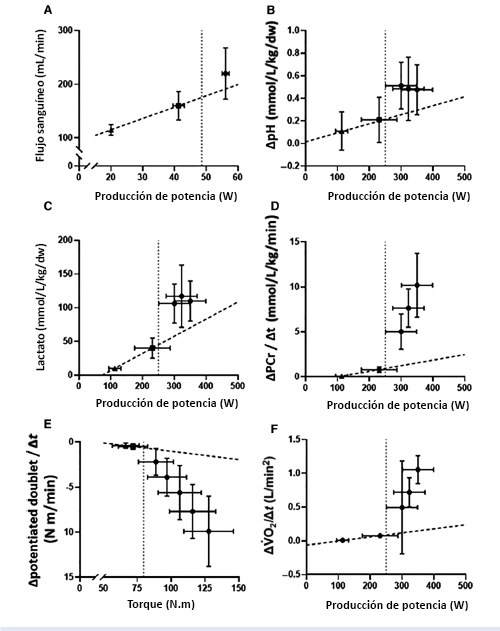
Figura 6. Valores expresados en forma de Media±SD del flujo sanguíneo muscular (Panel A; Copp et al. 2010), perturbación metabólica muscular (pH, Panel B; lactato, Panel C; Black et al. 2017) y tasas de cambio en la [PCr] muscular (Panel D; Black et al. 2017), excitabilidad neuromuscular (Panel E; Burnley et al. 2012) y VO2pulmonar(Panel F; Black et al. 2017) después del ejercicio de intensidad moderada (triángulos), de intensidad alta (cuadrados) y de intensidad severa (círculos).La línea vertical punteada indica la CP y se trazó la línea de mejor ajuste para todas las pruebas realizadas por debajo de CP (i.e.,ejercicio de intensidad moderada y alta;Línea discontinua).Es importante resaltar los cambios desproporcionados en todas las variables durante el ejercicio de intensidad severa (i.e., por encima de CP) en relación con el ejercicio realizado por debajo de CP.Estos datos revelan que la CP sería un umbral bioenergético por encima del cual se acelera el desarrollo de la fatiga y se impide la homeostasis muscular y sistémica.
Un argumento que se usa con frecuencia contra la idea de que la CP representa el límite entre el ejercicio de alta intensidad y el ejercicio de intensidad severa, es que la tolerancia al ejercicio no cumple con la idea de “tarea carente de fatiga” cuando se solicita a los participantes que realicen ejercicio de manera continua enla CP predeterminada (Poole et al. 1988; Jenkins y Quigley 1990; McLellan y Cheung 1992; Bull et al. 2000; Brickley et al. 2002; McClave et al. 2011; Bergstrom et al. 2013).Este argumento se basa en una interpretación errónea de la definición original de CP (Monod y Scherrer 1965) y es defectuoso, por dos razones.La primera es fundamental, ya que el modelo de potencia crítica de 2 parámetros no es aplicable para la predicción de la tolerancia al ejercicio precisamente en la CP (o por debajo de ella).De hecho, el límite tolerable (Tlim) de ejercicio en CP requeriría resolver la siguiente ecuación:
Tlim = W ‘/ (P – CP), donde P = CP, => Tlim = W’/0
Debido a que W/0 es matemáticamente falso, es ilógico juzgar la validez del modelo de CP sobre la base de una suposición de una “tarea sin fatiga” en CP.La segunda razón es metodológica, ya que realizar una prueba de ejercicio precisamente en CP no tiene en cuenta el error asociado con la estimación de CP. Si bien una ventaja de la determinación de la curva de duración-potencia es que permite la estimación de CP de hasta un solo watt , no es razonable considerar que este valor es absoluto. Los enfoques utilizados para modelar matemáticamente CP se asociarán naturalmente con algún error (que es cuantificable, por ejemplo, como error estándar o intervalos de confianza del 95%) y Tlim y CP variarán un poco en todos los individuos de un día a otro (es decir, existe una variabilidad biológica inherente; Poole et al., 1988). Por lo tanto, hay un “ancho de banda” o “área gris” que rodea a la CP estimada por el modelo, cuyo tamaño se puede minimizar a aproximadamente ± 3-5% con una cuidadosa atención al protocolo (ver más abajo). Por ejemplo, para una estimación de CP de 300 W y un error estándar de 2%, la CP “real” estará entre 294 y 306 W. Esto significa, sin embargo, que si este sujeto en particular realiza ejercicio a exactamente 300 W, entonces existe un 50% de probabilidad de que el/ella esté <CP y en el dominio de alta intensidad, y un 50% de probabilidad de que el/ella se ubique > CP y en el dominio de intensidad severa. Esto tendría implicaciones importantes para las respuestas fisiológicas, la naturaleza y la dinámica del desarrollo de la fatiga y de la tolerancia al ejercicio (Black et al. 2017). Por esta razón, no es sorprendente que cuando se solicita a los sujetos que realicen ejercicio en CP, el tiempo hasta el límite de tolerancia sea muy variable (por ejemplo, un rango de variación de aproximadamente 15 a 40 min u ocasionalmente hasta ~ 60 min; McLellan y Cheung 1992; Bull et al. 2000; Brickley et al. 2002; McClave et al. 2011; Bergstrom et al. 2013), y las respuestas fisiológicas medias del grupo serán características del ejercicio de alta intensidad (Poole et al. 1988, 1990; Wakayoshi et al. 1993) o del ejercicio de intensidad severa (Jenkins y Quigley 1990; McLellan y Cheung 1992; Brickley et al. 2002). Evidentemente, la práctica de solicitar a los sujetos que realicen ejercicios en la CP no es una prueba adecuada de la validez del concepto. De hecho, la cuestión misma de “cuánto tiempo” puede mantenerse la CP está mal concebida. El quid de la cuestión es que la CP separa un dominio de ejercicio dentro del cual se puede establecer la homeostasis fisiológica (dominio de alta intensidad) de otro dominio en el cual no se puede y en el cual la tolerancia al ejercicio es altamente predecible (dominio de intensidad severa).También se debe tener en cuenta que la duración del ejercicio en elMLSS nunca se ha determinado, pero esto también sería finalmente insostenible (Black et al. 2017).
La apreciación de la relación y de las diferencias entre el MLSS y la CP se ha confundido con la idea persistente pero desconcertante de que el máximo estado estable metabólico debería corresponder con una duración de ejercicio de aproximadamente 1 h. Esto es evidente suponiendo que el MLSS es equivalente a la llamada potencia “umbral funcional” que se puede mantener durante 60 minutos (Gavin et al. 2012; Morgan et al. 2018). Esta es una definición conveniente pero completamente arbitraria que carece de un significado fisiológico. No hay nada más “especial” en aproximadamente 60 minutos de ejercicio en comparación con, por ejemplo, 65 minutos, 44 minutos o 23 minutos. De hecho, el ejercicio máximo de 60 min de duración se posiciona directamente dentro del dominio de alta intensidad (Black et al. 2017), de manera que las respuestas fisiológicas al ejercicio máximo de una duración de 50-55 min o 65-70 min, probablemente serían muy similares en términos de los valores que se obtendrían al final del ejercicio y en la dinámica de respuesta. Un enfoque científico con una mayor justificación sería definir el máximo estado estable metabólico como la velocidad o la potencia que separa los distintos comportamientos de respuesta fisiológica, independientemente de la duración del ejercicio correspondiente. Es esperable que un enfoque de este tipo, que está anclado en el concepto de CP, pueda predecir mejor la capacidad de rendimiento y sea de mayor utilidad en la prescripción de ejercicios/entrenamiento (Jones et al. 2010; Vanhatalo et al. 2011a).
Es sorprendente que, cuando se conoce y se mide de manera apropiada el error estándar asociado a la estimación de la CP, las respuestas fisiológicas al ejercicio realizadas ligeramente por debajo y ligeramente por encima de la CP son esencialmente diferentes (Burnley et al. 2006; Jones et al. 2008; Murgatroyd et al. 2014; Vanhatalo et al. 2016). Cuando la CP se mide con cuidado, el error estándar asociado con la estimación del parámetro puede ser bastante pequeño (p. ej., 4 W en Vanhatalo et al. 2007) lo que genera confianza en que se puede determinar con precisión la transición auténtica entre un dominio de ejercicio en el que se podrá alcanzar la homeostasis (eventualmente) hacia un dominio en el que no se podrá alcanzar dicha homeostasis. Como se indicó anteriormente, esto es verdadero no solo para el intercambio de gases pulmonares, donde una característica fundamental del ejercicio de intensidad severa es el desarrollo de un componente lento del VO2que provocará que se alcance el VO2max en o cerca del punto de intolerancia al ejercicio (Poole et al. 1988; Hill et al. 2002; Vanhatalo et al. 2007; Jones et al. 2010), si no que también para la distribución del gasto cardíaco (Copp et al. 2010), para el equilibrio ácido-base de la sangre (Poole et al. 1998; Vanhatalo et al. 2016) y para los índices de metabolismo muscular (p. ej., [PCr] muscular y pH/lactato) ya sea que se determinen de forma no invasiva con 31 P-MRS (Jones et al. 2008) o de forma invasiva mediante biopsia (Vanhatalo et al. 2016; Black et al. 2017). La CP, por lo tanto, pasa (literalmente) la “prueba de acidez” de validez para separar el dominio de ejercicio de alta intensidad del de intensidad severa. Es importante enfatizar que no son solo los valores absolutos de las variables fisiológicas clave (p. ej.VO2, concentración de lactato sanguíneo, [PCr] muscular) determinadas en tiempos similares o el ejercicio final quienes permiten distinguir el ejercicio de intensidad severa del ejercicio de alta intensidad, si no que también las diferencias marcadas en los perfiles dinámicos de estas y otras variables (es decir, demora en el estado estable vs comportamiento de estado no estable).
El ejercicio realizado en diferentes potencias en el dominio de ejercicio de intensidad severa produce un estado metabólico muscular similar ([PCr], [Pi], pH, lactato) al que se observa en el límite de tolerancia (Vanhatalo et al. 2010; Black et al. 2017), consistente con la utilización de un W’ uniforme y finito, y con haber alcanzado el VO2max(Murgatroyd et al. 2011; Vanhatalo et al. 2011b) (Figura 7), mientras que estas variables no presentan el mismo grado de perturbación en el dominio de alta intensidad (ver Figura 6). Estos resultados indican que la PC permite diferenciar los dominios de intensidad del ejercicio dentro de los cuales predominan los diferentes mecanismos de desarrollo de fatiga (Black et al. 2017). De hecho, la evidencia disponible sugiere que la intolerancia al ejercicio está asociada con una contribución relativamente mayor de los sitios “periféricos” durante el ejercicio a realizados por encima de CP y una contribución relativamente mayor de los factores “centrales” junto con el agotamiento del glucógeno muscular durante los ejercicios realizados por debajo de CP (Burnley et al. 2012; Thomas et al. 2016; Black et al. 2017). Las diferencias en el desarrollo de fatiga durante los ejercicios realizados por debajo y por encima de la CP se analizan con más detalle en otros trabajos de investigación (Poole et al. 2016; Burnley y Jones 2018). Sin embargo, es pertinente reiterar aquí que es posible que los sujetos realicen ejercicios por encima del MLSS y todavía produzcan respuestas fisiológicas consistentes con el ejercicio de alta intensidad (por ejemplo, Mattioni Maturana et al. 2016), lo que sugiere que el MLSS no separa precisamente los dominios de intensidad del ejercicio donde los perfiles de respuesta fisiológica y los mecanismos de desarrollo de la fatiga son diferentes.
Consideraciones para la Evaluación Precisa de la Relación Potencia-Duración
Aunque hemos revisado la evidencia que apoya a la CP como elparámetro delimitador de buena fedel máximo estado estable metabólico, es fundamental que se tenga mucho cuidado en su estimación (Mattioni Maturana et al. 2018; Muniz-Pumares et al. 2019). Existen dos métodos mediante los cuales se puede evaluar la CP: la metodología “convencional” en la cual la CP se modela a partir de una serie de “pruebas de predicción” de intensidad severa realizadas hasta el límite de tolerancia en diferentes velocidades o producciones de potencia (Monod y Scherrer 1965; Poole et al. 1988); y el test máximo de 3 minutos en donde, como su nombre lo indica, los sujetos realizan ejercicios al máximo durante 3 minutos y en donde la potencia de la prueba final es equivalente a la CP y el trabajo total realizado por encima de la CP es equivalente a W’ (Burnley et al. 2006; Vanhatalo et al. 2007). Si la CP se estima utilizando el enfoque convencional, es importante considerar el número de pruebas y su duración (Hill 1993; Bishop et al. 1998; Triska et al. 2018). Es esencial que los sujetos hagan su máximo esfuerzo en cada prueba y que la cadencia sea consistente en todas las pruebas. Idealmente, la prueba más corta debería ser de 2-3 minutos y la más larga debería ser superior a 10 min pero no más de 15 min (Hill 1993; Vanhatalo et al. 2011a). Es recomendable que exista al menos una diferencia de 5 minutos entre las pruebas más cortas y las más largas (Bishop et al. 1998) pero la bondad del ajuste hiperbólico aumenta al hacer el rango de tiempo hasta el agotamiento lo más amplio posible (i.e., 8-12 min) dentro del dominio de intensidad severa. La duración precisa de las pruebas de predicción tiene una importancia secundaria para el alcance del VO2máx, pero es inusual que se alcance VO2máxsi la duración del ejercicio es inferior a 1-2 minutos o superior a 15-20 min (Hill et al. 2002; Vanhatalo et al. 2016). El VO2 debe ser medido durante cada prueba para verificar que se alcance el VO2máxel cual se define típicamente como el VO2del final del ejercicio quesupera el 95% del VO2máxmedido durante el ejercicio incremental en rampas, para permitir la variabilidad biológica y metodológica diaria (Katch et al. 1982).
La bondad de ajuste de los datos de la prueba de predicción con la ecuación de regresión depende del número de pruebas. En la práctica, 3-4 (Smith y Jones 2001; Brickley et al. 2002; Pringle y Jones 2002; Dekerle et al. 2005; Black et al. 2015) o 5-7 (Hughson et al. 1984; Gaesser y Wilson 1988 Poole et al., 1990; Bull et al., 2000; Vanhatalo et al., 2007). La bondad del ajuste, que se informa en forma de valores r2, nos brinda solamente una indicación amplia de precisión. Se recomienda aportar los errores estándar (o intervalos de confianza del 95%) asociados con cada parámetro, con una precisión que se considere satisfactoria cuando el error estándar sea inferior al 5% de la media para CP e inferior al 10% para el W (Hill y Smith 1994, 1999). Los datos de las pruebas de predicción deben ser modelados de forma iterativa y es necesario realizar pruebas adicionales hasta que se alcancen estos criterios de SE. La variabilidad natural en el rendimiento de resistencia humano de una prueba a otra significa que las estimaciones de los parámetros que se obtienen de los tres modelos de 2 parámetros para medir (es decir, el modelo lineal de tiempo de trabajo, el modelo hiperbólico de tiempo-potencia y el modelo lineal 1/tiempo; Hill 1993; Morton 2006) rara vez son idénticas. La aplicación de los tres modelos y la búsqueda del “mejor ajuste individual” (es decir, el modelo que produce el menor error combinado para CP y W) para cada sujeto es un enfoque útil (Black et al. 2015, 2017). La variabilidad en el rendimiento también puede dar como resultado pequeñas diferencias en la CP estimada cuando se combina un pequeño número de pruebas de predicción, incluso cuando todas las pruebas están dentro del rango recomendado (por ejemplo, 3, 7 y 12 minutos) vs. 2, 5 y 10 min; Triska et al. 2018). Es esperable que existan diferencias menores dado que las pruebas de predicción discretas de diferentes duraciones solo pueden aportar una aproximación del continuo fundamental potencia-tiempo subyacente; tales diferencias no socavan la validez de la CP, sino que resaltan la importancia de utilizar estrategias apropiadas para minimizar el error de medición.
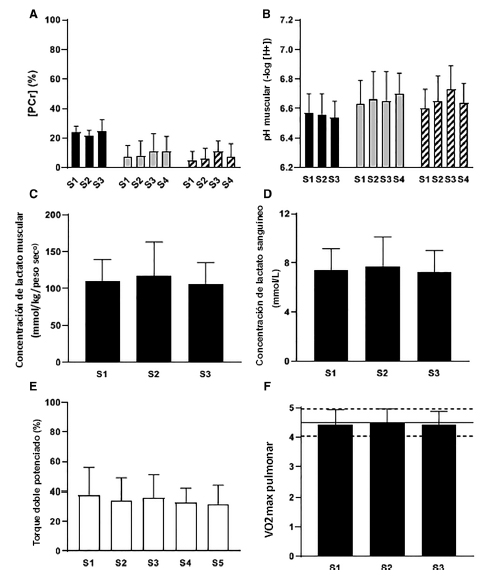
Figura 7. Losparticipantes que realizan ejercicio de intensidad severa alcanzan el mismo medio metabólico muscular “crítico” ([PCr] panel A, pH en el Panel B y concentración de lactato sanguíneo en Panel C); presentan valores de concentración de lactato sanguíneo similares (Panel D); experimentan disminuciones equivalentes en la excitabilidad neuromuscular (Panel E); y alcanzan el VO2max pulmonar (Panel F), en el límite de tolerancia independientemente de la duración del ejercicio. Estas respuestas se observan después de ejercicios de ciclismo (barras negras, Black et al. 2017) y de ejercicios de extensión de la rodilla realizados en normoxia (barras blancas, Burnley et al. 2012; barras gris claro, Vanhatalo et al. 2010) e hiperoxia (70 % O2 , barras gris oscuro, Vanhatalo et al. 2010). Se presentan los valores grupales en forma de Media±SD. La línea continua en el Panel F, indica el VO2max determinado a partir del test incremental en rampas. S1 = serie de ejercicios de intensidad severa 1, y así sucesivamente.
La estimación de la CP se puede acelerar utilizando el test máximo de 3 minutos, que se realiza pedaleando contra una resistencia fija en una bicicleta ergométrica Lode Excalibur Sporto en una pista, y se ha demostrado que aporta información válida y estimaciones confiables de CP (Burnley et al. 2006; Vanhatalo et al. 2007, 2008a; Pettitt et al. 2012; Broxterman et al. 2013; Simpson et al. 2015). Es importante que los sujetos estén altamente motivados y completamente familiarizados con el protocolo en su totalidad, y comprendan que deben realizar un esfuerzo máximo durante toda la prueba. Es necesario tener mucho cuidado en la normalización de la resistencia fija para realizar la prueba de ciclismo en la bicicleta ergométrica. Las variables que se miden en el test máximo de 3 minutos son sensibles a la manipulación de la cadencia (Vanhatalo et al. 2008b; Wright et al. 2019), por lo que la selección de una “cadencia preferida” que sea demasiado alta (≥ 90 rpm) tiende a provocar una subestimación de W´ y una sobreestimación de CP. El profesional que supervise la prueba no debe proporcionar retroalimentación sobre el tiempo durante la prueba y el estímulo verbal debe ser constante para garantizar que se realice con la misma urgencia y entusiasmo en todo momento. Una cantidad sustancial de evidencia indica que, en sujetos recreativamente activos, el VO2max en el test máximo de 3 minutos por lo general alcanza aprox 97-103% del VO2max establecido en el test con rampas (Burnley et al. 2006; Vanhatalo et al. 2011b; Barker et al. 2012; Chidnok et al. 2013; Black et al. 2015; Clark et al. 2018). Por lo tanto, para que la prueba de criterio sea aceptada como válida, no debe haber indicación de mantener un ritmo en el perfil de velocidad o producción de potencia (es decir, no debe existir una tendencia incremental en la producción de potencia o en la velocidad en ningún punto después de la aceleración inicial durante los primeros 5-10 segundos), y el VO2maxdebe ser alcanzado y luego mantenido durante el resto de la prueba (Jones et al. 2010; Vanhatalo et al. 2016). Si el VO2 obtenido durante una prueba máxima de 3 minutos es <95% VO2max, las estimaciones de CP y W´ no deben ser consideradas como precisas y la prueba debe ser repetida.
Aplicación Práctica de la Relación Potencia-Duración
La cuantificación de la relación potencia-duración utilizando los procedimientos de prueba descriptos previamente nos aportará no solo el valor de CP (asíntota de la relación) sino también el valor de W´ (la constante de la curva de la relación).La CP es un índice de capacidad metabólica oxidativa que es sensible al entrenamiento de resistencia (Gaesser y Wilson 1988; Poole et al. 1990; Jenkins y Quigley 1992; Vanhatalo et al. 2008a) y a la fracción de O2 inspirado (Vanhatalo et al. 2010; Dekerle et al. 2012; Simpson et al. 2015 La Mónica et al. 2018). La CP está altamente correlacionada con el rendimiento de los ejercicios de resistencia (Kolbe et al. 1995; Smith et al. 1999; Black et al. 2014) y se ha estimado que los corredores de maratón de élite mantienen aproximadamente 96% de su velocidad crítica durante las competencias (Jones y Vanhatalo 2017). Por otra parte y algo importante, el W´ aporta información sobre la cantidad finita de trabajo que se puede realizar durante un ejercicio que se realiza por encima de la CP antes de alcanzar el límite de tolerancia (Fukuba et al. 2003; Chidnok et al. 2013) y es sensible a las intervenciones que alteran el componente lento del VO2 (Vanhatalo et al. 2010; Murgatroyd et al. 2011). Durante el ejercicio de intensidad severa, que contempla eventos de resistencia en el rango de aprox. 2-25 min, el rendimiento es una función de la interacción de CP con W´ (Vanhatalo et al. 2011a; Jones y Vanhatalo 2017). Por lo tanto, aunque la CP sola nos aporta información sobre la mas alta tasa metabólica oxidativa sostenible durante el ejercicio de alta intensidad, conocer tanto la CP como el W´ permite una predicción altamente precisa del rendimiento durante el ejercicio de intensidad severa (Vanhatalo et al. 2011a; Morgan et al. 2018) y es muy valioso para la confección de programas de entrenamiento intervalado optimizados individualmente (Skiba et al. 2014; Jones y Vanhatalo 2017). Sin embargo, es necesario destacar que tanto CP como W´ son cantidades dinámicas que pueden disminuir con el tiempo durante ejercicios extenuantes (Clark et al. 2018, 2019).
Conclusiones
El concepto de máximo estado estable metabólico es valioso desde múltiples perspectivas, como mejorar nuestra comprensión de la energía básica del músculo esquelético y los procesos de fatiga, para caracterizar la intensidad del ejercicio y para explorar y superar las limitaciones para el rendimiento durante el ejercicio humano. Sin embargo, el progreso en estos campos se ha retrasado, debido a la falta de acuerdo sobre las definiciones y los procedimientos, y por una fijación con el comportamiento de un marcador biológico único, la concentración de lactato sanguíneo. En este artículo, hemos esbozado las preocupaciones sobre la arbitrariedad de la definición y de los procedimientos para evaluar el MLSS, y hemos proporcionado una justificación para considerar a la CP como el límite que separa al ejercicio en estado estable (alta intensidad) del ejercicio en estado no estable (intensidad severa). Recomendamos a los científicos y profesionales que puedan evidenciar que MLSS y CP no son sinónimos intercambiables y no debe esperarse que lo sean. Las diferencias cuantitativas y cualitativas entre estas entidades son inevitables, y son causadas por la definición conservadora de MLSS que produce una subestimación del límite de intensidad alta/severa establecido en función de la CP. Como todas las otras mediciones en fisiología humana (ejercicio), existe un error técnico obligatorio y una variabilidad biológica inherente a la estimación de la CP. Sin embargo, cuando estos factores se minimizan mediante un procedimiento experimental sólido, y se cuantifican y explican adecuadamente, es evidente que la CP permite separar los dominios de intensidad del ejercicio con perfiles distintivos de respuestas cardiovasculares, respiratorias metabólicas y sistémicas. Por lo tanto, la CP es el parámetro adecuado cuando el objetivo es evaluar el máximo estado estable metabólico.
Conflicto de Intereses
Los autores no poseen ningún conflicto de interés.
Para citar este artículo en su versión original
Andrew M. Jones, Mark Burnley, Matthew I. Black, David C. Poole and Anni Vanhatalo (2019). The maximal metabolic steady state: redefining the “gold standard”. Physiol. Rep, 7 (10), 2019, e14098. doi: 10.14814/phy2.14098
Referencias
Barker, T., D. C. Poole, M. L. Noble, and T. J. Barstow. 2006. Human critical power-oxygen uptake relationship at different pedalling frequencies. Exp. Physiol. 91:621–632.
Barker, A. R., B. Bond, C. Toman, C. A. Williams, and N. Armstrong. 2012. Critical power in adolescents: physiological bases and assessment using all-out exercise. Eur. J. Appl. Physiol. 112:1359–1370.
Beneke, R. 1995. Anaerobic threshold, individual anaerobic threshold, and maximal lactate steady state in rowing. Med. Sci. Sports Exerc. 27:863–867.
Beneke, R. 2003. Methodological aspects of maximal lactate steady state-implications for performance testing. Eur. J. Appl. Physiol. 89:95–99.
Beneke, R., and S. P. von Duvillard. 1996. Determination of maximal lactate steady state response in selected sports events. Med. Sci. Sports Exerc. 28:241–246.
Beneke, R., M. H€utler, and R. M. Leith€auser. 2000. Maximal lactate-steady-state independent of performance. Med. Sci. Sports Exerc. 32:1135–1139.
Beneke, R., R. M. Leith€auser, and M. H€utler. 2001. Dependence of the maximal lactate steady state on the motor pattern of exercise. Br. J. Sports Med. 35:192–196.
Beneke, R., M. Hutler, S. P. Von Duvillard, M. Sellens, and R. M. Leithauser. 2003. Effect of test interruptions on blood lactate during constant workload testing. Med. Sci. Sports Exerc. 35:1626–1630.
Bergman, B. C., E. E. Wolfel, G. E. Butterfield, G. D. Lopaschuk, G. A. Casazza, M. A. Horning, et al. 1999.
Active muscle and whole body lactate kinetics after endurance training in men. J. Appl. Physiol. 87:1684–1896.
Bergstrom, H. C., T. J. Housh, J. M. Zuniga, D. A. Traylor, R. W. Lewis, C. L. Camic, et al. 2013. Responses during exhaustive exercise at critical power determined from the 3-minute all-out test. J. Sports Sci. 31:537–545.
Billat, V., F. Dalmay, M. T. Antonini, and A. P. Chassain. 1994. A method for determining the maximal steady state of blood lactate concentration from two levels of submaximal exercise. Eur. J. Appl. Physiol. Occup. Physiol. 69:196–202.
Billat, V. L., P. Sirvent, G. Py, J. P. Koralsztein, and J. Mercier. 2003. The concept of maximal lactate steady state: a bridge between biochemistry, physiology and sport science. Sports Med. 33:407–426.
Billat, V. L., E. Mouisel, N. Roblot, and J. Melki. 2005. Interand intrastrain variation in mouse critical running speed. J. Appl. Physiol. 98:1258–1263.
Bishop, D., D. G. Jenkins, and A. Howard. 1998. The critical power function is dependent on the duration of the
predictive exercise tests chosen. Int. J. Sports Med. 19:125– 129.
Black, M. I., J. Durant, A. M. Jones, and A. Vanhatalo. 2014.
Critical power derived from a 3-minute all-out test predicts 16.1-km road time-trial performance. Eur. J. Sport Sci. 14:217–223.
Black, M. I., A. M. Jones, S. J. Bailey, and A. Vanhatalo. 2015. Self-pacing increases critical power and improves
performance during severe-intensity exercise. Appl. Physiol. Nutr. Metab. 40:662–670.
Black, M. I., A. M. Jones, J. R. Blackwell, S. J. Bailey, L. J. Wylie, S. T. McDonagh, et al. 2017. Muscle metabolic and neuromuscular determinants of fatigue during cycling in different exercise intensity domains. J. Appl. Physiol. 122:446–459.
Bonaventura, J. M., K. Sharpe, E. Knight, K. L. Fuller, R. K. Tanner, and C. J. Gore. 2015. Reliability and accuracy of six hand-held blood lactate analysers. J. Sports Sci. Med.
14:203–214.
Brickley, G., J. Doust, and C. A. Williams. 2002. Physiological responses during exercise to exhaustion at critical power. Eur. J. Appl. Physiol. 88:146–151.
Broxterman, R. M., C. J. Ade, D. C. Poole, C. A. Harms, and T. J. Barstow. 2013. A single test for the determination of parameters of the speed-time relationship for running. Respir. Physiol. Neurobiol. 185:380–385.
Bull, A. J., T. J. Housh, G. O. Johnson, and S. R. Perry. 2000. Effect of mathematical modeling on the estimation of critical power. Med. Sci. Sports Exerc. 32:526–530.
Burnley, M. 2009. Estimation of critical torque using intermittent isometric maximal voluntary contractions of
the quadriceps in humans. J. Appl. Physiol. 106:975–983.
Burnley, M., and A. M. Jones. 2018. Power-duration relationship: Physiology, fatigue, and the limits of human
performance. Eur. J. Sport Sci. 18:1–12.
Burnley, M., J. H. Doust, and A. Vanhatalo. 2006. A 3-minute all-out test to determine peak oxygen uptake and the maximal steady state. Med. Sci. Sports Exerc. 38:1995–2003.
Burnley, M., A. Vanhatalo, and A. M. Jones. 2012. Distinct profiles of neuromuscular fatigue during muscle
contractions below and above the critical torque in humans. J. Appl. Physiol. 113:215–223.
Carter, H., A. M. Jones, and J. H. Doust. 1999. Effect of 6 weeks of endurance training on the lactate minimum
speed. J. Sports Sci. 17:957–967.
Carter, H., J. S. Pringle, A. M. Jones, and J. H. Doust. 2002. Oxygen uptake kinetics during treadmill running across exercise intensity domains. Eur. J. Appl. Physiol. 86:347–354.
Chidnok, W., F. J. Dimenna, S. J. Bailey, D. P. Wilkerson, A. Vanhatalo, and A. M. Jones. 2013. Effects of pacing strategy on work done above critical power during highintensity exercise. Med. Sci. Sports Exerc. 45:1377–1385.
Clark, I. E., A. Vanhatalo, S. J. Bailey, L. J. Wylie, B. S. Kirby, B. W. Wilkins, et al. 2018. Effects of two hours of heavyintensity exercise on the power-duration relationship. Med. Sci. Sports Exerc. 50:1658–1668.
Clark, I, A. Vanhatalo, C. Thompson, L. J. Wylie, S. J. Bailey, B. Kirby, et al. 2019. Changes in the power-duration relationship following prolonged exercise: estimation using conventional and all-out protocols and relationship to muscle glycogen. Am. J. Physiol. Regul. Integr. Comp. Physiol.. https://doi.org/10.1152/ajpregu.00031.2019. [Epubahead of print]\
Copp, S. W., D. M. Hirai, T. I. Musch, and D. C. Poole. 2010. Critical speed in the rat: implications for hindlimb muscle blood flow distribution and fibre recruitment. J. Physiol. 588:5077–5087.
Davies, M. J., A. P. Benson, D. T. Cannon, S. Marwood, G. J. Kemp, H. B. Rossiter, et al. 2017. Dissociating external power from intramuscular exercise intensity during intermittent bilateral knee-extension in humans. J. Physiol. 595:6673–6686.
Dekerle, J., B. Baron, L. Dupont, J. Vanvelcenaher, and P. Pelayo. 2003. Maximal lactate steady state, respiratory
compensation threshold and critical power. Eur. J. Appl. Physiol. 89:281–288.
Dekerle, J., P. Pelayo, B. Clipet, S. Depretz, T. Lefevre, and M. Sidney. 2005. Critical swimming speed does not represent the speed at maximal lactate steady state. Int. J. Sports Med.
26:524–530.
Dekerle, J., P. Mucci, and H. Carter. 2012. Influence of moderate hypoxia on tolerance to high-intensity exercise.
Eur. J. Appl. Physiol. 112:327–335.
Dill, D. B., and D. L. Costill. 1974. Calculation of percentage changes in volumes of blood, plasma, and red cells in dehydration. J. Appl. Physiol. 37:247–248.
Faude, O., W. Kindermann, and T. Meyer. 2009. Lactate threshold concepts: how valid are they? Sports Med.
39:469–490.
Fukuba, Y., A. Miura, M. Endo, A. Kan, K. Yanagawa, and B. J. Whipp. 2003. The curvature constant parameter of the power-duration curve for varied-power exercise. Med. Sci. Sports Exerc. 35:1413–1418.
Full, R. J. 1986. Locomotion without lungs: energetics and
performance of a lungless salamander. Am. J. Physiol. 251: R775–R780.
Full, R. J., and C. F. 2nd Herreid. 1983. Aerobic response to exercise of the fastest land crab. Am. J. Physiol. 244:R530–R536.
Gaesser, G. A., and L. A. Wilson. 1988. Effects of continuous and interval training on the parameters of the powerendurance time relationship for high-intensity exercise. Int. J. Sports Med. 9:417–421.
Gaesser, G. A., S. A. Ward, V. C. Baum, and B. J. Whipp. 1994. Effects of infused epinephrine on slow phase of O2 uptake kinetics during heavy exercise in humans. J. Appl. Physiol. 77:2413–2419.
Gavin, T. P., J. B. Van Meter, P. M. Brophy, G. S. Dubis, K. N. Potts, and R. C. Hickner. 2012. Comparison of a field-based test to estimate functional threshold power and power output at lactate threshold. J. Strength Cond. Res. 26:416–421.
Heck, H., A. Mader, G. Hess, S. M€ucke, R. M€uller, and W. Hollmann. 1985. Justification of the 4-mmol/l lactate threshold. Int. J. Sports Med. 6:117–130.
Hermansen, L., E. Hultman, and B. Saltin. 1967. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 71:129–139.
Hill, A. V. 1925. The physiological basis of athletic records. Nature 116:544–548.
Hill, D. W. 1993. The critical power concept. A review. Sports Med. 16:237–254.
Hill, D. W., and C. S. Ferguson. 1999. A physiological description of critical velocity. Eur. J. Appl. Physiol. Occup. Physiol. 79:290–293.
Hill, D. W., and J. C. Smith. 1994. A method to ensure accuracy of estimates of anaerobic capacity derived using
the critical power concept. J. Sports Med. Physical Fitness. 34:23–37.
Hill, D. W., and J. C. Smith. 1999. Determination of critical power by pulmonary gas exchange. Can. J. Appl. Physiol. 24:74–86.
Hill, D. W., D. C. Poole, and J. C. Smith. 2002. The relationship between power and the time to achieve VO
(2max). Med. Sci. Sports Exerc. 34:709–714.
Holloszy, J. O., and E. F. Coyle. 1984. Adaptations of skeletal muscle to endurance exercise and their metabolic
consequences. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 56:831–838.
Hughson, R. L., C. J. Orok, and L. E. Staudt. 1984. A high velocity treadmill running test to assess endurance running potential. Int. J. Sports Med. 5:23–25.
Iannetta, D., E. C. Inglis, C. Fullerton, L. Passfield, and J. M. Murias. 2018. Metabolic and performance-related
consequences of exercising at and slightly above MLSS. Scand. J. Med. Sci. Sports 28:2481–2493.
Jacobs, I. 1986. Blood lactate. Implications for training and sports performance. Sports Med. 3:10–25.
Jamnick, N. A., J. Botella, D. B. Pyne, and D. J. Bishop. 2018.Manipulating graded exercise test variables affects the validity of the lactate threshold and VO2 peak. PLoS ONE
Jenkins, D. G., and B. M. Quigley. 1990. Blood lactate in trained cyclists during cycle ergometry at critical power. Eur. J. Appl. Physiol. Occup. Physiol. 61:278–283.
Jenkins, D. G., and B. M. Quigley. 1992. Endurance training enhances critical power. Med. Sci. Sports Exerc. 24:1283–1289.
Jones, A. M., and H. Carter. 2000. The effect of endurance training on parameters of aerobic fitness. Sports Med.
29:373–386.
Jones, A. M., and J. H. Doust. 1998. The validity of the lactate minimum test for determination of the maximal lactate steady state. Med. Sci. Sports Exerc. 30:1304–1313.
Jones, A. M., and D. C. Poole. 2008. Physiological demands of endurance exercise. Pp. 43–55 in R. J. Maughan, (Ed). Olympic Textbook of Science in Sport, IOC, Blackwell, Oxford, UK.
Jones, A. M., and A. Vanhatalo. 2017. The ‘critical power’ concept: applications to sports performance with a focus on intermittent high-intensity exercise. Sports Med. 47:65–78.
Jones, A. M., D. P. Wilkerson, F. DiMenna, J. Fulford, and D. C. Poole. 2008. Muscle metabolic responses to exercise above and below the “critical power” assessed using 31P-MRS. Am. J. Physiol. Regul. Integr. Comp. Physiol. 294:R585–R593.
Jones, A. M., A. Vanhatalo, M. Burnley, R. H. Morton, and D. C. Poole. 2010. Critical power: implications for
determination of VO2max and exercise tolerance. Med. Sci. Sports Exerc. 42:1876–1890.
Jones, A. M., B. Grassi, P. M. Christensen, P. Krustrup, J. Bangsbo, and D. C. Poole. 2011. Slow component of VO2 kinetics: mechanistic bases and practical applications. Med. Sci. Sports Exerc. 43:2046–2062.
Jones, A. M., M. Burnley, and A. Vanhatalo. 2018. Aerobic Exercise Performance. Pp. 319–352 In K. Norton, R. Eston, eds. Kinanthropometry and Exercise Physiology. Routledge, London. https://doi.org/10.4324/9781315385662
Jorfeldt, L., A. Juhlin-Dannfelt, and J. Karlsson. 1978. Lactate release in relation to tissue lactate in human skeletal muscle during exercise. J. Appl. Physiol. Respir. Environ. Exerc. Physiol. 44:350–352.
Katch, V. L., S. S. Sady, and P. Freedson. 1982. Biological variability in maximum aerobic power. Med. Sci. Sports Exerc. 14:21–25.
Keir, D. A., F. Y. Fontana, T. C. Robertson, J. M. Murias, D. H. Paterson, J. M. Kowalchuk, et al. 2015. Exercise intensity thresholds: identifying the boundaries of sustainable performance. Med. Sci. Sports Exerc. 47:1932–1340.
Kilding, A. E., and A. M. Jones. 2005. Validity of a single-visit protocol to estimate the maximum lactate steady state. Med. Sci. Sports Exerc. 37:1734–1740.
Kolbe, T., S. C. Dennis, E. Selley, T. D. Noakes, and M. I. Lambert. 1995. The relationship between critical power and running performance. J. Sports Sci. 13:265–269.
La Monica, M. B., D. H. Fukuda, T. M. Starling-Smith, R. Wang, J. R. Hoffman, and J. R. Stout. 2018. Effects of normobaric hypoxia on upper body critical power and anaerobic working capacity. Respir. Physiol. Neurobiol.
249:1–6.
LaFontaine, T. P., B. R. Londeree, and W. K. Spath. 1981. The maximal steady state versus selected running events. Med. Sci. Sports Exerc. 13:190–193.
Lauderdale, M. A., and K. W. Hinchcliff. 1999. Hyperbolic relationship between time-to-fatigue and workload. Equine Vet. J. Suppl. 30:586–590.
Mader, A., and H. Heck. 1986. A theory of the metabolic origin of anaerobic threshold. Int. J. Sports Med. 7:45–65.
Mattioni Maturana, F., D. A. Keir, K. M. McLay, and J. M. Murias. 2016. Can measures of critical power precisely estimate the maximal metabolic steady-state? Appl. Physiol. Nutr. Metab. 41:1197–1203.
Mattioni Maturana, F., F. Y. Fontana, S. Pogliaghi, L. Passfield, and J. M. Murias. 2018. Critical power: How different protocols and models affect its determination. J. Sci. Med. Sport. 21:742–747.
McClave, S. A.,M. LeBlanc, and S. A.Hawkins. 2011. Sustainability of critical power determined by a 3-minuteute all-out test in elite cyclists. J. Strength Cond. Res. 25:3093–3098.
McLellan, T. M., and K. S. Cheung. 1992. A comparative evaluation of the individual anaerobic threshold and the
critical power. Med. Sci. Sports Exerc. 24:543–550.
Mitchell, E. A., N. R. W. Martin, S. J. Bailey, and R. A. Ferguson. 2018. Critical power is positively related to skeletal muscle capillarity and type I muscle fibers in endurance-trained individuals. J. Appl. Physiol. 125:737–745.
Monod, H., and J. Scherrer. 1965. The work capacity of a synergic muscular group. Ergonomics 8:329–338.
Morgan, P. T., M. I. Black, S. J. Bailey, A. M. Jones, and A. Vanhatalo. 2018. Road cycle TT performance: Relationship to the power-duration model and association with FTP. J. Sports Sci. 37:902–910. Nov 2:1-9. https://doi.org/10.1080/02640414.2018.1535772.
Moritani, T., A. Nagata, H. A. deVries, and M. Muro. 1981. Critical power as a measure of physical work capacity and anaerobic threshold. Ergonomics 24:339–350.
Morton, R. H. 2006. The critical power and related wholebody bioenergetic models. Eur. J. Appl. Physiol. 96:339–354.
Muniz-Pumares, D., B. Karsten, C. Triska, and M. Glaister. 2019. Methodological approaches and related challenges associated with the determination of critical power and curvature constant. J. Strength Cond. Res. 33:584–596.
Murgatroyd, S. R., C. Ferguson, S. A. Ward, B. J. Whipp, and H. B. Rossiter. 2011. Pulmonary O2 uptake kinetics as a determinant of high-intensity exercise tolerance in humans. J. Appl. Physiol. 110:1598–1606.
Murgatroyd, S. R., L. A. Wylde, D. T. Cannon, S. A. Ward, and amd H. B. Rossiter. 2014. A ‘ramp-sprint’ protocol to characterise indices of aerobic function and exercise intensity domains in a single laboratory test. Eur.
Pettitt, R. W., N. Jamnick, and I. E. Clark. 2012. 3-minute allout exercise test for running. Int. J. Sports Med. 33:426–431.
Poole, D. C., and R. S. Richardson. 1997. Determinants ofoxygen uptake. Implications for exercise testing. Sports
Med. 24:308–320.
Poole, D. C., S. A. Ward, G. W. Gardner, and B. J. Whipp. 1988. Metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics 31:1265–1279.
Poole, D. C., S. A. Ward, and B. J. Whipp. 1990. The effects of training on the metabolic and respiratory profile of highintensity cycle ergometer exercise. Eur. J. Appl. Physiol. Occup. Physiol. 59:421–429.
Poole, D. C., M. Burnley, A. Vanhatalo, H. B. Rossiter, and A. M. Jones. 2016. Critical power: an important fatigue threshold in exercise physiology. Med. Sci. Sports Exerc. 48:2320–2334.
Priest, J. W., and R. D. Hagan. 1987. The effects of maximum steady state pace training on running performance. Br. J. Sports Med. 21:18–21.
Pringle, J. S., and A. M. Jones. 2002. Maximal lactate steady state, critical power and EMG during cycling. Eur. J. Appl. Physiol. 88:214–226.
Saunders, P. U., D. B. Pyne, R. D. Telford, and J. A. Hawley. 2004. Reliability and variability of running economy in elite distance runners. Med. Sci. Sports Exerc. 36:1972–1976.
Scheen, A., J. Juchmes, and A. Cession-Fossion. 1981. Critical analysis of the “anaerobic threshold” during exercise at constant workloads. Eur. J. Appl. Physiol. Occup. Physiol. 46:367–377.
Simpson, L. P., A. M. Jones, P. F. Skiba, A. Vanhatalo, and D. Wilkerson. 2015. Influence of hypoxia on the powerduration relationship during high-intensity exercise. Int. J. Sports Med. 36:113–119.
Sj€odin, B., I. Jacobs, and J. Svedenhag. 1982. Changes in onset of blood lactate accumulation (OBLA) and muscle enzymes after training at OBLA. Eur. J. Appl. Physiol. Occup. Physiol. 49:45–57.
Skiba, P. F., D. Clarke, A. Vanhatalo, and A. M. Jones. 2014. Validation of a novel intermittent W’ model for cycling using field data. Int. J. Sports Physiol. Perform. 9:900–904.
Smith, C. G., and A. M. Jones. 2001. The relationship between critical velocity, maximal lactate steady-state velocity and lactate turnpoint velocity in runners. Eur. J. Appl. Physiol. 85:19–26.
Smith, J. C., B. S. Dangelmaier, and D. W. Hill. 1999. Critical power is related to cycling time trial performance. Int. J. Sports Med. 20:374–378.
Snyder, A. C., T. Woulfe, R. Welsh, and C. Foster. 1994. A simplified approach to estimating the maximal lactate steady state. Int. J. Sports Med. 15:27–31.
Stainsby, W. N., and G. A. Brooks. 1990. Control of lactic acid metabolism in contracting muscles and during exercise. Exerc. Sport Sci. Rev. 18:29–63.
Stegmann, H., and W. Kindermann. 1982. Comparison of prolonged exercise tests at the individual anaerobic threshold and the fixed anaerobic threshold of 4 mmol.l(-1) lactate. Int. J. Sports Med. 3:105–110.
Tesch, P. A., W. L. Daniels, and D. S. Sharp. 1982. Lactate accumulation in muscle and blood during submaximal
exercise. Acta Physiol. Scand. 114:441–446.
Thomas, K., M. Elmeua, G. Howatson, and S. Goodall. 2016. Intensity-dependent contribution of neuromuscular fatigue after constant-load cycling. Med. Sci. Sports Exerc. 48:1751–1760.
Triska, C., B. Karsten, C. Beedie, B. Koller-Zeisler, A. Nimmerichter, and H. Tschan. 2018. Different durations within the method of best practice affect the parameters of the speed-duration relationship. Eur. J. Sport Sci. 18:332–340.
Vanhatalo, A., J. H. Doust, and M. Burnley. 2007. Determination of critical power using a 3-minute all-out
cycling test. Med. Sci. Sports Exerc. 39:548–555.
Vanhatalo, A., J. H. Doust, and M. Burnley. 2008a. A 3- minute all-out cycling test is sensitive to a change in critical power. Med. Sci. Sports Exerc. 40:1693–1699.
Vanhatalo, A., J. H. Doust, and M. Burnley. 2008b. Robustness of a 3 min all-out cycling test to manipulations of power profile and cadence in humans. Exp. Physiol. 93:383–390.
Vanhatalo, A., J. Fulford, F. J. DiMenna, and A. M. Jones. 2010. Influence of hyperoxia on muscle metabolic responses and the power-duration relationship during severe-intensity exercise in humans: a 31P magnetic resonance spectroscopy study. Exp. Physiol. 95:528–540.
Vanhatalo, A., A. M. Jones, and M. Burnley. 2011a. Application of critical power in sport. Int. J. Sports Physiol.
Perform. 6:128–136.
Vanhatalo, A., D. C. Poole, F. J. DiMenna, S. J. Bailey, and A. M. Jones. 2011b. Muscle fiber recruitment and the slow component of O2 uptake: constant work rate vs. all-out sprint exercise. Am. J. Physiol. Regul. Integr. Comp. Physiol. 300:R700–R707.
Vanhatalo, A., M. I. Black, F. J. DiMenna, J. R. Blackwell, J. F. Schmidt, C. Thompson, et al. 2016. The mechanistic bases of the power-time relationship: muscle metabolic responses and relationships to muscle fibre type. J. Physiol. 594:4407–4423.
Wakayoshi, K., T. Yoshida, M. Udo, T. Harada, T. Moritani, Y. Mutoh, et al. 1993. Does critical swimming velocity represent exercise intensity at maximal lactate steady state? Eur. J. Appl. Physiol. Occup. Physiol. 66:90–95.
Whipp, B. J., and S. A. Ward. 1992. Pulmonary gas exchange dynamics and the tolerance to muscular exercise: effects of fitness and training. Ann. Physiol. Anthropol. 11:207–214.
Wilkerson, D. P., K. Koppo, T. J. Barstow, and A. M. Jones. 2004. Effect of work rate on the functional ‘gain’ of Phase II pulmonary O2 uptake response to exercise. Respir. Physiol. Neurobiol. 142:211–223.
Wilkie, D. R. 1960. Man as a source of mechanical power. Ergonomics 3:1–8.
Womack, C. J., S. E. Davis, J. L. Blumer, E. Barrett, A. L. Weltman, and G. A. Gaesser. 1995. Slow component of O2 uptake during heavy exercise: adaptation to endurance training. J. Appl. Physiol. 79:838–845.
Wright, J., S. Bruce-Low, and S. A. Jobson. 2019. The 3- minuteute all-out cycling test is sensitive to changes in
cadence using the Lode Excalibur Sport ergometer. J. Sports Sci. 37:156–162.
Yamamoto, Y., M. Miyashita, R. L. Hughson, S. Tamura, M. Shinohara, and Y. Mutoh. 1991. The ventilatory threshold gives maximal lactate steady state. Eur. J. Appl. Physiol. Occup. Physiol. 63:55–59.