
04 Nov Transportadores de Lactato
Antes de comenzar con el desarrollo del término en cuestión, vemos conveniente aclarar una serie de
conceptos relacionados con éste, y así comprender mejor ha este grupo de proteínas, su función y el papel que pueden desempeñar en el contexto del entrenamiento deportivo.
El término “shuttle” o lanzadera atiende a funciones de transporte de ciertas moléculas o substancias de un lugar hacia otro. En el caso que nos atiende, estamos hablando del transporte del lactato y por consiguiente de sus vías metabólicas.
El transporte de lactato es llevado a cabo por una familia de proteínas de transporte monocarboxiladas (MCTs), que se expresan en células y tejidos gracias a sus múltiples isoformas (en
tiempos actuales se describen hasta 14 tipos existentes (Ortega Diez, 2014)). Por ejemplo, en el tejido cardíaco se ha identificado la isoforma MCT1, en el musculo esquelético las isoformas MCT1/MCT4; entre muchas otras descubiertas en distintos tipos de tejidos (Bonen, 2000).
deshidrogenasa mitocondrial (LDHm) una vez en el interior de la mitocondria donde podrá ser oxidado (San Millán, 2014). Otro ejemplo de lanzadera intramuscular es la que produce el intercambio de piruvato por lactato causado en los peroxisomas (Brooks, 2009). La segunda lanzadera es llamada de tipo célula a célula, que como su nombre indica es aquella donde se produce un flujo de lactato entre distintas células de un mismo o diferente tejido y/o órgano. Algunos ejemplos (figura 1) de lanzaderas célula-célula incluyen el intercambio de lactato entre las fibras glucolíticas (blancas) y oxidativas (rojas) dentro de un músculo que está ejercitándose, pero también entre el músculo esquelético que trabaja y el corazón, el cerebro, el hígado o los riñones(Brooks, 2009). En esta definición vamos a profundizar un poco más sobre la lanzadera de lactato célula a célula.
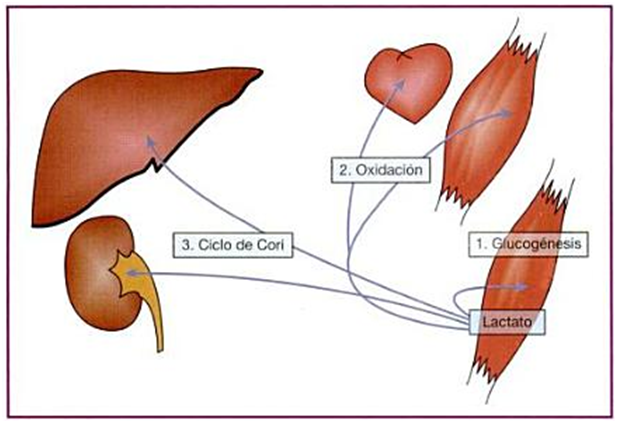
Figura 1. Ejemplos del destino e intercambio del lactato producido a nivel muscular gracias al
sistema de lanzaderas a través de las distintas isoformas de MCT. 1: precursor gluconeogénico en el propio musculo; 2: sustrato para ser oxidado en otros músculos (incluyendo el miocardio) y 3: formación de glucógeno en el ciclo de Cori. (Extraído de Chicharro&Vaquero, 2006).
A modo de síntesis podemos decir que el lactato es un producto glucolítico que puede ser utilizado tanto en (a) las células en las que se originó, (b) transportado a través del espacio intersticial hacia
otras células, o (c) a través de la sangre hacia otros tejidos. De esta manera el lactato es un sustrato oxidable importante, además de que gracias a su distribución tiene la capacidad de coordinar el metabolismo en diversos tejidos (Fernanda, 2003).
Este último punto obedece a la teoría de la lanzadera de lactato (Brooks, 1985), donde se presupone que el lactato es removido a tejidos anatómica e histológicamente iguales o diferentes, a distancia (célula a célula), brindando una fuente significativa de sustrato oxidable y de precursor gluconeogénico.
El transporte del lactato a través del sarcolema muscular (membrana externa que recubre la célula muscular) se produce por tres vías (Chicharro&Vaquero, 2006) las cuáles se describen a continuación:
1) MCTs, la cual es la manera mayoritaria en que se moviliza el lactato (y la cual desarrollamos con este término) y funciona a través de un sistema de cotransporte junto a un protón (figura 2), lo cual es de gran importancia para la regulación del pH, especialmente durante la actividad muscular intensa (Juel, 2001).
2) Un intercambiador aniónico que intercambia el anión lactato por Cl- o HCO3- .
3) Y por último, la difusión de ácido láctico no disociado.
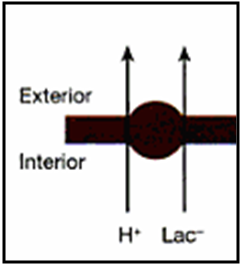
Figura 2. Mecanismo de transporte de lactato hacia fuera la membrana muscular mediante la proteínas transportadoras monocarboxiladas (MCT). Este mecanismo, juntamente a otros, es además un sistema de regulación del pH a nivel muscular (Modificado de Chicharro&Vaquero, 2006).
Las isoformas musculares de los MCT mencionados anteriormente, tienen características propias y funciones diferenciadas. El MCT1 está más relacionado con el papel energético que el lactato puede
proporcionar como sustrato en la célula muscular, por lo tanto, está vinculado a la captación de éste a su interior para su posterior oxidación, sin embargo, el MCT4 está más relacionado con la capacidad glucolítica y participa en la eliminación (salida) del lactato de la célula muscular (Chicharro&Vaquero, 2006). Así entonces, vista la evidencia, Bonen y colaboradores (2000) sugieren que los MCT1 son los principales responsables de la captación de lactato de la circulación (y también de la lanzadera intramuscular, anteriormente mencionada, al introducir el lactato en las mitocondrias una vez dentro la célula) y los MCT4 de la salida del lactato muscular.
Dadas estas peculiaridades, parece lógico pensar que las fibras con mayor carácter oxidativo presenten una concentración mayor de MCT1 que de MCT4, lo contrario que sus pares glucolíticas (Bonen, 2000). Esto se confirmó al ver que el transportador MCT1 ha sido encontrado de forma predominante en fibras musculares mayormente oxidativas y sólo en pequeñas cantidades en las glucolíticas, sin embargo, el MCT4 se expresa en forma significativa en los dos tipos de fibras (Fernanda, 2003). Es importante no olvidar este último punto para comprender lo mencionado a continuación.
Aunque parezca que vaya en contra de la lógica mencionada hasta ahora, se ha visto que el transporte de lactato es significativamente más rápido en la fibras oxidativas que en sus pares glucolíticos (Juel, 2001). Vale la penar recordar, que si bien las fibras oxidativas tienen un alto potencial de captación y oxidación del lactato, también son proclives a su formación y liberación; además, al estar usualmente más tiempo bajo ejercitación que las glucolíticas, el tener un transporte de lactato más rápido les daría una ventaja fisiológica. Éste puede ser uno de los motivos del porqué este tipo de fibras presentan una mayor resistencia a la fatiga al compararse con las glucolíticas,
que al tener un transporte de lactato hacia fuera la célula más lento, ésta pudiera ser la razón del porque después de un ejercicio intenso, el lactato postejercicio retenido en ellas sea resintetizado en forma de glucógeno (Fernanda, 2003) como se ha comentado anteriormente.
fuerte medida el metabolismo láctico. Respecto este último enfoque más práctico, nos gustaría enlazar el término con un artículo muy interesante que se publicó hace relativamente poco aquí en nuestro portal, el cual habla sobre una propuesta de entrenamiento intervalado basada en el control por frecuencia cardíaca aplicada en nadadores, donde tuvo efectos positivos en los mecanismos de transporte de lactato mencionados a lo largo del término.
Autor
David Masferrer Llana
Referencias
Bonen, A., 2000. Lactate transporters (MCT proteins) in heart and skeletal muscles: Med. Sci. Sports Exerc. 32, 778–789.
Brooks, G.A., 1985. Lactate:Glycolytic End Product and Oxidative Substrate During Sustained Exercise in Mammals — The “Lactate Shuttle,” in: Gilles, P.D.R. (Ed.), Circulation, Respiration, and Metabolism, Proceedings in Life Sciences. Springer Berlin Heidelberg, pp. 208–218.
Brooks, G.A., 2009. Cell–cell and intracellular lactate shuttles. J. Physiol. 587, 5591–5600.
Brooks, G.A., Dubouchaud, H., Brown, M., Sicurello, J.P., Butz, C.E., 1999. Role of mitochondrial lactate dehydrogenase and lactate oxidation in the intracellular lactate shuttle. Proc. Natl. Acad. Sci. 96, 1129–1134.
Chicharro, J.L., Vaquero, A.F., 2006. Fisiologa del ejercicio / Physiology of Exercise. Ed. Médica Panamericana.
Juel, C., 2001. Current aspects of lactate exchange: lactate/H+ transport in human skeletal muscle. Eur. J. Appl. Physiol. 86, 12–16.
Ortega Diez, J., 2014. Series de Frecuencia Cardiaca o “Series de Repeticiones Australianas.” PubliCE Lite.
Pilegaard, H., Domino, K., Noland, T., Juel, C., Hellsten, Y., Halestrap, A.P., Bangsbo, J., 1999. Effect of high-intensity exercise training on lactate/H+ transport capacity in human skeletal muscle. Am. J. Physiol. – Endocrinol. Metab. 276, E255–E261.